Long Covid, Short Magnesium - Chambers
Preprint and published in April 2022
the following is extracted from the preprint
📄 Click Here for the published PDF Scientific Research Publishing
Abstract
April 2022 DOI: 10.13140/RG.2.2.31726.08009
Patrick W Chambers
Long Covid, now also reported in children, has become increasingly alarming. Perhaps a third of those who develop MIS-C go on to experience Long Covid. Etiology is elusive. The role of vitamin D in bone and immune health has been recognized but magnesium deficiency has escaped attention. A physiologic and biochemical argument for its culpability in Long Covid as well Chronic Fatigue Syndrome, Fibromyalgia, Epstein Barr Virus, and Cytomegalovirus is presented.
📄 Download the PDF from VitaminDWiki
Introduction
The link between vitamin D deficiency and rickets was first discovered in the 1920s. It was not until 50 years later, in the 1970s, that the entity, magnesium deficient, vitamin D resistant rickets, was realized. In 1971 Jean Durlach founded the Society for the Development and Research on Magnesium (SDRM). But despite two excellent books on the topic, The Magnesium Factor (2003) by Mildred Seelig/Andrea Rosanoff and The Magnesium Miracle (2006 first ed., 2017 second ed.) by Carolyn Dean, research has languished.
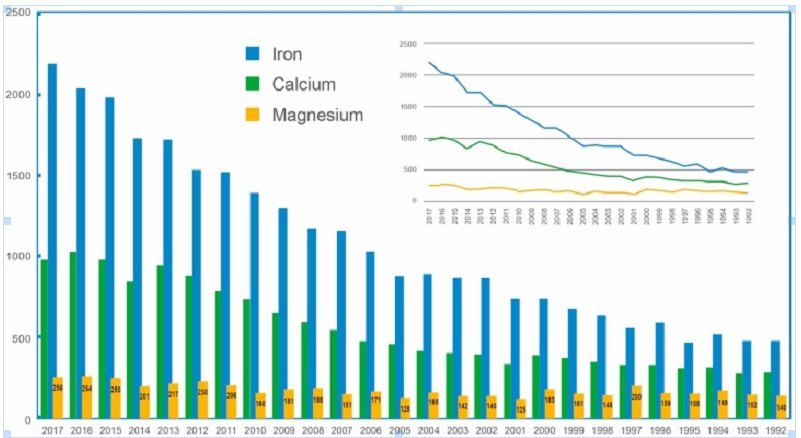
Figure 1 . The number of basic and clinical research papers published (Y-axis) on Fe, Ca, and Mg 1 .
Magnesium metabolism and requirements are inextricably entwined with vitamin D. One cannot discuss vitamin D without discussing magnesium. Similarly, one cannot correct a vitamin D deficiency associated problem without magnesium sufficiency.
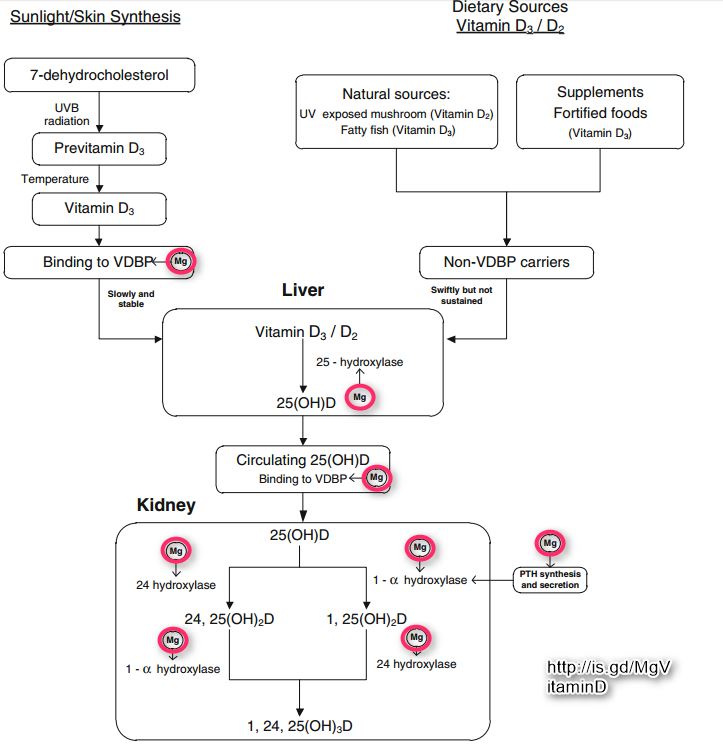
Figure 2. Magnesium is not only required for seven critical steps in the synthesis of active vitamin D 1,25(OH){SUB()}2{SUB}D but is also required for the synthesis and secretion of PTH by the parathyroid (and other endocrine) glands.
However, sudden increases of vitamin D in the presence of magnesium deficiency is problematic. Existing free Mg2+ could be triaged from low priority functions e.g., neuromuscular, to servicing the added D3/25(OH)D3. The result could be symptoms of magnesium deficiency, e.g., worsening constipation or increase in palpitations. Repleting D alone will deplete Mg2+. This has tarnished the value of vitamin D supplementation in many who are also magnesium deficient. In many the double deficiency persists and future supplementation shunned.
Hypothesis
Magnesium deficiency is intimately involved in the brain fog, fatigue, post-exertional malaise, headache, dizziness, myalgia and asthma of Long Covid or Post Acute Coronavirus Syndrome (PACS).
Discussion
What Objectively Constitutes Magnesium Deficiency?
A recent study demonstrated that dietary magnesium is inversely related to serum parathormone (PTH) concentration in the overweight and obese, especially in response to raising serum 25(OH)D concentration 2 .
Based on dietary magnesium intake, three groups over-weight or obese (n=57) were identified: low Mg intake group (median 131 mg/day); intermediate intake group (median 216.9 mg/day); high intake group (median 376 mg/day). (RDA for magnesium is 310 to 320 for women and 400 to 420 for men). The magnesium intake in the group reporting benefit (lower PTH) approached that recommended by the RDA. The median body mass index (BMI) for all three groups was 27.7-29, close to the median American BMI of 26.5.
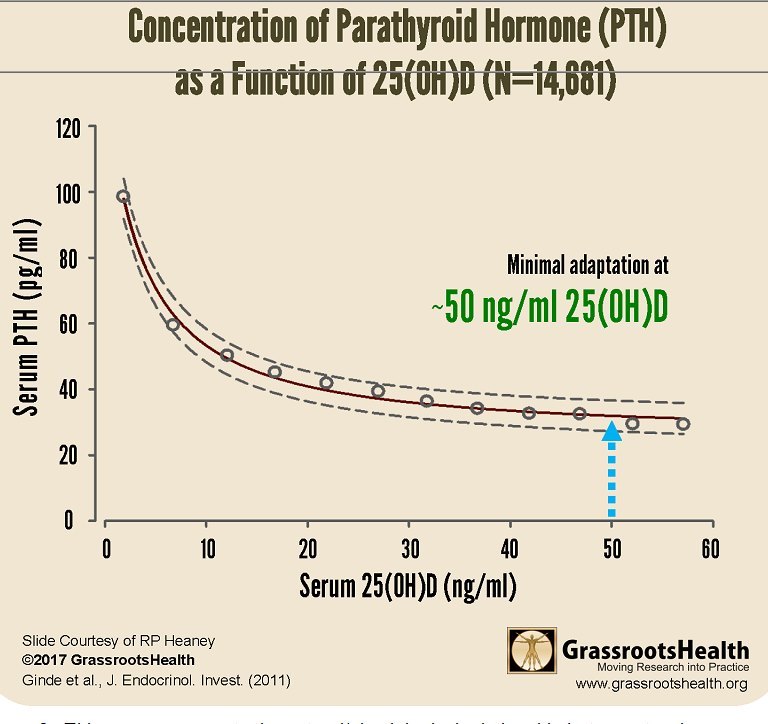
Figure 3. This curve represents the natural/physiological relationship between two hormones with plateauing of the curve when serum 25(OH)D concentration is approximately 50 ng/mL.
The study subjects in the three groups had 25(OH)D levels between 24.6 and 27.1 ng/mL, i.e., all were magnesium deficient. However, according to Figure 3, any value less than 50 ng/mL would have shown similar results, since the PTH/25(OH)D relationship is still suboptimal below that amount. Gastrointestinal and renal absorption of magnesium is influenced by vitamin D, which is controlled by PTH. The latter is predominantly controlled by ionised calcium Ca2+ and Mg2+ to a lesser extent.
The flattening of the curve at serum 25(OH)D concentrations above 50 ng/mL seems to imply an optimum balance between calcium, magnesium, PTH (and calcitonin) above this set point. In the study serum 25(OH)D concentration was inversely associated with PTH only in the high Mg2+ intake group. This latter has several implications.
1) The lower magnesium intake is entirely consumed by bone needs without significant change in blood Ca or PTH.
2) In the high intake group not all the magnesium is consumed by bone needs. The surplus is redirected from bone to needs for vitamin D synthesis (Figure 2). Note: At least in the renal tubular cells, PTH enhances the conversion of 25(OH)D into 1,25(OH){SUB()}2{SUB}D (magnesium is a required cofactor), which in turn decreases PTH secretion (Figure 2).
At the lower intakes, all magnesium is consumed by bone needs, since the PTH was already elevated (indicating the presence of some degree of secondary hyperparathyroidism and perhaps, overactive osteoclastic activity), with no magnesium left to synthesize 1,25(OH){SUB()}2{SUB}D to address the osteoporosis. We know that everyone in the study is vitamin D deficient (< 30 ng/mL). But this study shows that they are still deficient at anything less than 50 ng/mL. The study also demonstrates that magnesium is deficient at this set point. Vitamin D requirement of the obese is 2 to 3 times greater than that for those of normal weight 3 .
One can appreciate the importance of magnesium and the complexity of its physiology, especially when known and unknown feedback loops, polymorphisms (genes), nutrition, lifestyles, etc., are considered. Randomized clinical trials (RCTs) are rarely comparable; some of them incorporate controversial assumptions, are expensive, and are time-consuming.
This 50 ng/mL is not a blood serum concentration level recommended by most meta-analyses or some prospective NIH-funded-study, but one ordained by Mother Nature through human evolution. The production of vitamin D3 (cholecalciferol) in the skin from sunlight is essentially limited to the four hours between 10 am and 2 pm. This benefit is also seasonal and needs are directly proportional to BMI. Generally, with the current lifestyles, solar exposure is rarely sufficient and supplementation rarely adequate. Both routes to the active form are magnesium-dependent (Figure 2).
Once the minimum 50 ng/mL vitamin D level has been established, evaluating magnesium status becomes feasible. Unfortunately, biologic individuality, e.,g., genes or single nucleotide polymorphisms (SNPs), and the complex nature of measuring free intracellular Mg2+ make accurate evaluation difficult. There are no laboratory tests that are reliable in this regard. This makes a bottom-up approach superior to a top-down one.
NOTE
Magnesium supplementation is fraught with difficulty. Most have a laxative effect, especially magnesium citrate. Exceeding bowel tolerance results in loss of magnesium and potassium. Magnesium in chelated form to malate, threonate, glycinate, or another amino acid is recommended. Aqueous magnesium (Mg2+) was previously considered highly absorbable, but the size of the hydrated cation compromises passive ion channel passage, which is responsible for the vast majority of small bowel absorption see figures 4,5 at 1. Furthermore absorption via active transport requires ATP and is magnesium dependent. It is also important to differentiate the magnesium content by weight per tablet (elemental Mg or Mg as …) versus total tablet weight and to note how many tablets per serving. Magnesium (MgCl{SUB()}2{SUB}, MgSO{SUB()}4{SUB}) can also be absorbed through hair follicles.
What Is the Triage Theory of Vitamins/Minerals and Does It Apply to Vitamin D/Magnesium?
In 2006 Bruce Ames4 posited that when micronutrient availability is limited, those functions required for short-term survival take precedence over those less urgent. In the case of magnesium, it appears that bone needs for magnesium supersede those needed to produce active vitamin D. After all vitamin D deficient rickets was described in the 1920s, but magnesium-deficient vitamin D-resistant magnesium rickets was not recognized until the 1970s.
The recommended 30 ng/mL of 25(OH)D is sufficient to avoid rickets in the presence of magnesium sufficiency but is inadequate to avoid secondary hyperparathyroidism and osteoporosis. Furthermore, the minimum magnesium intake for any individual should be that necessary to maintain a 25(OH)D level above 50 ng/mlmL. Nevertheless, there is no measure estimating whether this amount of magnesium is sufficient to address magnesium needs for a healthy immune system.
Based on the higher incidence of osteoporosis and the lower incidence of COVID-19 in Northern Europe versus the opposite in Southern Europe, any magnesium shortfall is triaged first to bone needs. In second place are Vitamin D synthesis needs, which supersede those of immunomodulation. The high dietary Ca and 25(OH)D levels in Northern Europe create a magnesium shortfall by suppressing PTH. PTH down-regulates vitamin D, decreasing magnesium intestinal absorption and renal resorption, decreasing blood levels of magnesium (innocent bystander) and aggravating osteoporosis. Clearly, adequate vitamin D is more critical to immunomodulation than adequate magnesium, given the low incidence of Covid-19 in Northern Europe.
Their Southern cousins, on the other hand, have sufficient magnesium to avoid osteoporosis, but not enough to avoid a greater risk of Covid-19. Their 25(OH)D3 levels are lower and SARS CoV2 symptomatology is more often severe. There is no significant osteoporosis => vitamin D, calcium, and magnesium needs are in balance, i.e., healthy bone needs for calcium and magnesium have been satisfied, as have magnesium needs for synthesis of active vitamin D. However, there is a problem with immunomodulation compared to their northern neighbors. This confirms that magnesium needs for immunomodulation are subservient to magnesium needs for healthy bone and vitamin D synthesis.
Since neuromuscular needs are probably in the runt position, we can only assure sufficient magnesium to fulfill any immunomodulation needs by addressing neuromuscular symptoms often associated with insufficiency, via magnesium supplementation. The contribution of magnesium to immune function independent of vitamin D is vastly under appreciated.
1. Why is Magnesium Deficiency Increasing?
It is decreasing because the quality of our food has decreased (Figure 4).
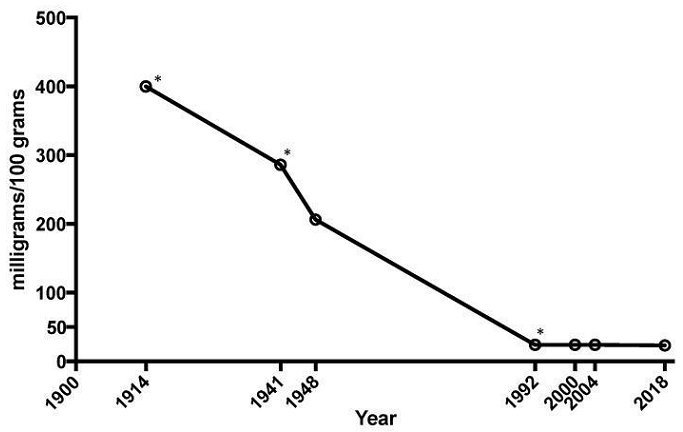
Figure 4. The average mineral content of calcium, magnesium, and iron in cabbage, lettuce, tomatoes, and spinach has dropped 80–90% between 1914 and 20181.
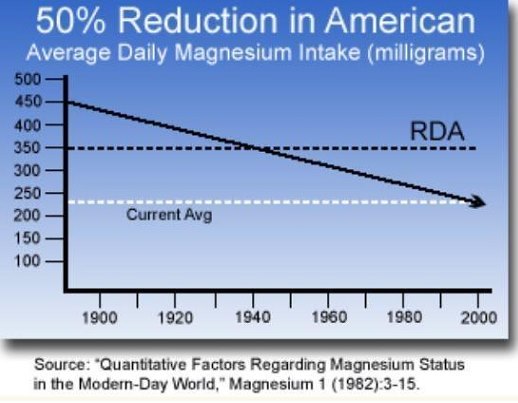
Figure 5. This graph indicates the need for magnesium supplementation.
Furthermore, an increasingly sedentary lifestyle, now with work at home via Zoom, precludes adequate sun exposure, aggravating many health issues and compromising vitamin D status. Inflammatory conditions deplete the already low stores of other vital vitamins and minerals, e.g., selenium, vitamin C, zinc,….
Humans in Western countries are consuming four times more calcium than magnesium versus the recommended 2:1 ratio (Figure 5). This is one of the reasons why osteoporosis is so high in those on diets high in dairy. The pharmaceutical industry has been conveniently available to address any symptoms due to a magnesium shortfall and/or any associated pain and suffering with a growing menu of helpful medications. Many of these exacerbate magnesium deficiency.
Dehydration is at the center of muscle cramps and headaches, commonly seen in magnesium deficiency. Aldosterone triggers renal reabsorption of Na+ and water, but at the expense of K+ and Mg++, which are excreted to maintain electrical neutrality. Stress is also a major player. The hormone cortisol from the adrenal cortex exerts aldosterone-like activity.
1. What Is the Role of Magnesium Outside Bone Health and Vitamin D Synthesis?
Magnesium is a critical mineral in the human body and is involved in ~80% of known metabolic functions. It is currently estimated that 60% of adults do not achieve the average dietary intake (ADI) and 45% of Americans are magnesium deficient ===1=== . Magnesium is a required cofactor for about 600 enzymatic reactions (many require vitamin B6) and serves as an activator for perhaps an additional 2005 (many if not all of these involve ATP).
The most significant of these is its role (as an activator) storing and releasing energy generated by ATP (Figure 6). In green plants it is also involved in generating ATP through chlorophyll in photosynthesis. That’s why eating leafy greens is healthful.
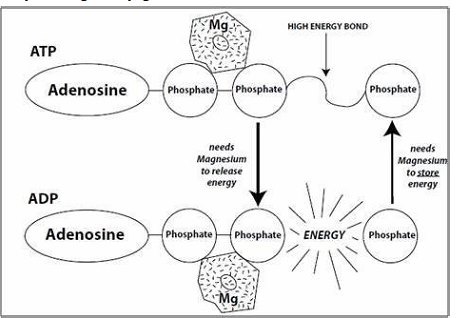
Figure 6. ATP and energy generation cannot proceed in the absence of magnesium (from The Magnesium Factor by Seelig and Rosanoff).
Magnesium is vital to the conduction of electrical impulses in nerves and muscles. There are three types of muscles - smooth, skeletal, and cardiac. Manifestations of magnesium deficiency in smooth muscle include constipation, bronchial asthma, and vasospasms (migraine headaches and even Prinzmetal angina due to coronary vasospasm). In skeletal muscle, it’s cramps and twitches (fasciculations). In heart muscle it’s palpitations (premature atrial contractions) and even atrial fibrillation. Any of these symptoms should put a magnesium shortfall on the radar.
In the immune system, the role of vitamin D is well known but poorly embraced. The flu shot has only 10% of the efficacy of an adequately maintained vitamin D. Magnesium (independent of vitamin D) is required for many disparate functions. But the role of magnesium in immune function is poorly understood and relatively unexplored.
Some well known natural antioxidants, e.g., glutathione and melatonin, require a magnesium cofactor for their synthesis. Melatonin, which decreases with age, also balances Th1 and Th2 cytokine responses, mimicking the benefits of vitamin D6. However, the role of magnesium as a receptor activator via ATP in immune function is especially intriguing. These are called receptor tyrosine kinases (RTKs) and include all the interferon receptors7. These ATP/Mg dependent RTKs include JAKs and TYK2 (Janus kinases 1,2,3, tyrosine kinase 2), which spotlights a very pertinent role for magnesium in immunomodulation.
1. Might Any Particular Cytokine Connect Magnesium Deficiency to Long Covid?
Secretion of type I interferons (INF-α, IFN-β) and type III interferons (IFN-λ) are innate immune cytokines (early) induced directly by invading viruses (primary release). At the same time, type II interferon gamma (IFN-γ) is predominantly an adaptive immune cytokine (secondary release), induced by NK (natural killer) cells (innate) and CD4+/CD8+ T cells (adaptive) via IFNGRs (IFN-γ receptors)8.
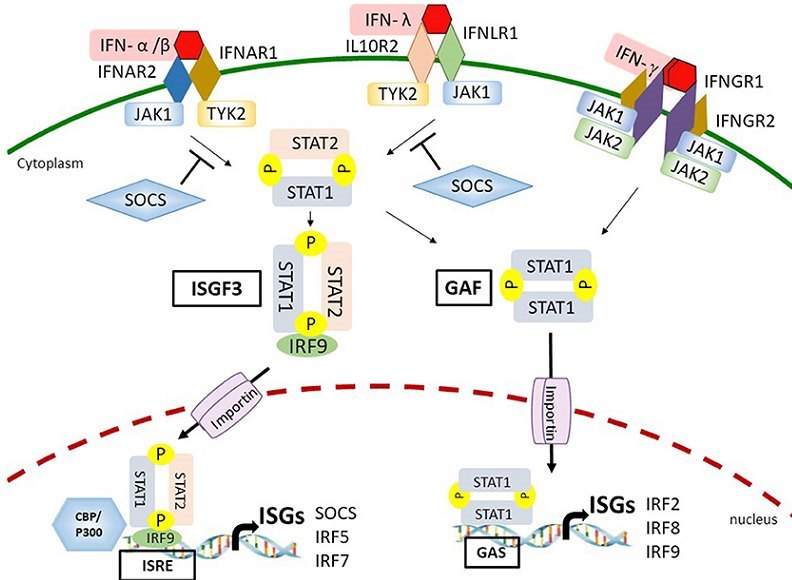
Figure 7. This demonstrates the secondary release of interferons. Note the yellow Ps, which indicate ATP/Mg dependency. JAK1,2 and TYK2 are also similarly dependent9 (P not shown).
JAKs1,2, TYK2, STATs1,2 are all ATP/Mg dependent. So one can appreciate the overwhelming role of ATP/Mg in interferon production alone. A magnesium shortfall might contribute to a decrease in IFN-γ and adaptive immunity. IFN-γ in particular is fundamental to understanding the symptoms of Long Covid and an alphabet soup of chronic inflammatory conditions (ME/CFS, FM, EBV, CMV) and autoimmune diseases (MS, SLE, RA, DM, …). These NK cells and CD4+/CD8+ T cells are the very PBMCs (peripheral blood mononuclear cells) that are exhausted during prolonged COVID-19 (lymphopenia). Plasma levels of IFN-γ were significantly reduced in Covid ICU patients8 , ===10=== , ===11=== . IFN-γ is a Th1 pro-inflammatory cytokine. But it has several beneficial effects, including blocking differentiation of a naive CD4+ T (Th0) cell into a Th17 cell and enhancing synthesis of the inhibitor of C1 of the CCP (Classic Complement Pathway).
1. What Causes Long Covid?
The diagnosis of Long Covid or PACS has become a bit of a mixed bag that can include those that never recovered from the primary infection, those that were never infected but developed similar symptoms during the prolonged quarantining and social isolation, those that recovered but developed unrelated but attributed symptoms, …. This complicates rational analysis. Accordingly discussion is directed primarily toward those outpatient/hospitalized for Covid that developed cognitive dysfunction or brain fog.
PACS or Long Covid exhibits a unique immunoglobulin signature (decreased IgM and IgG3)12. Ironically IgG3 and IFN-γ are proportionately elevated in both mild and severe COVID-19 cases13. IFNγ actually induces IgG3 secretion14. Only IgG1 and IgG3 can activate CCP and only the CCP crosstalks with the KKS. Long Covid can be predicted in those with indeterminate (low) IGRA (IFN-γ release assay) results15, ===16=== . Low IgG3 levels have also been linked to chronic fatigue syndrome (CFS)17,18, which suggests IFNγ may also be lower in CFS. IFNγ enhances synthesis of C1-INH in PBMCs and the liver19, ===20=== , suppressing over-activation of the CCP. So low IgG3 and low IFNγ in Long Covid and CFS translate to less C1-INH and more C1 activation. Once C1 has been activated, the CCP begins crosstalk with the KKS (Kallikrein Kinin System)21.
The lectin complement pathway (LCP) and the alternative complement pathway do not cross talk with the KKS.
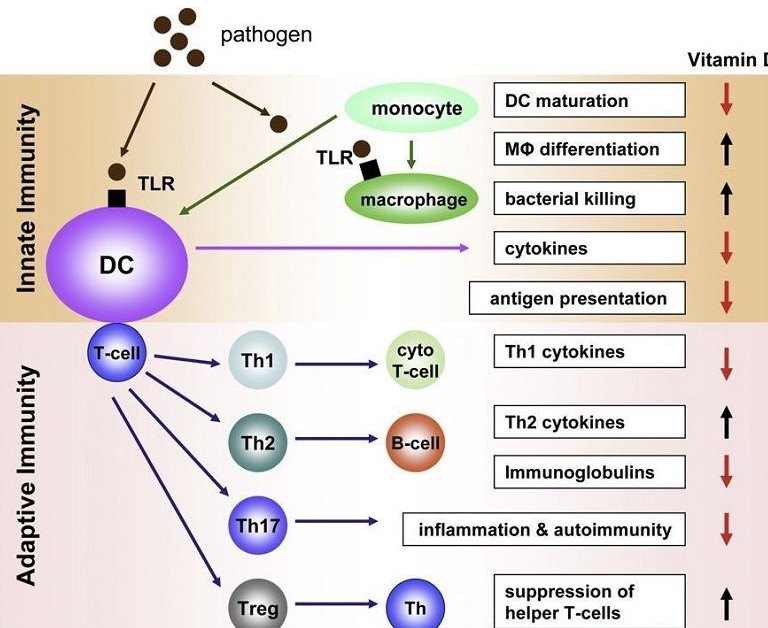
Figure 8. Vitamin D operates in both the innate and adaptive immunity phases, promotes anti-inflammatory cytokines, and balances the Th1/Th2 immune response22.
There are two types of asthma, allergic and non-allergic. Vitamin D was adequate **23** D%20>%2030%20ng/mL)%20in%20only%2015%%20of%20pediatric%20asthmatics%20v%2080%%20of%20healthy%20controls"http://dx.doi.org/10.2147/JAA.S29566"). This suggests that the majority of asthmatics are non-allergic. The risk of severe disease in COVID-19 is increased in non-allergic asthma[ 24,25. On the other hand allergic asthma (defined by the presence of eosinophilia) provides protection against COVID-19(26). Might much of this non-allergic asthma be due to magnesium deficiency causing bronchial smooth muscle spasm(27)? Might this group of asthmatics be at greater risk of Long Covid (and MIS-C)(12)?
This is the same pediatric sub-population (asthmatic and vitamin D deficient) that develops MIS-C. So, asthma in these children and adults (MIS-A) may be a marker for magnesium deficiency. Unfortunately in those at risk for MIS-C (and Long Covid) a sudden increase in vitamin D intake without simultaneously adding magnesium could further exacerbate symptoms due to triage. In addition to vitamin D, melatonin also helps balance the Th1/Th2 response6. But magnesium is a required cofactor for the synthesis of serotonin, the precursor to melatonin. Melatonin and asthma seem to connect magnesium to Long Covid.
Low IFNγ in Covid-19 is also associated with lung fibrosis28 => post exertional malaise, breathlessness, fatigue in Long Covid. Toxoplasma gondii, Cryptosporidium, Blastocyst, and Giardia infections downgraded Covid-19 severity via elevation of IFNγ29. This speaks to the beneficial C1 inhibiting properties of IFNγ, i.e., inhibition of CCP with inhibition of crosstalk to the KKS. Therefore, depleted IFNγ seems to translate to an acquired angioedema type clinical condition. Long Covid brain fog, fatigue, and breathlessness may be a function of low IFNγ, causing lung fibrosis and less C1 INH (more C1) with activation of the CCP and crosstalk with the KKS.
Due to the extensive ATP/Mg requirements for interferon release, magnesium deficiency could easily result in low IFNγ levels/low melatonin levels and mimic or exacerbate Long Covid30. Indeed one could probably expand this to include CFS, FM, EBV (chronic phase), and CMV (chronic phase) in combination with magnesium deficiency. All vitamin D receptors (VDRs) require zinc. Zinc deficiency could also masquerade as vitamin D deficiency and Long Covid31.
MIS-C32 (and probably Kawasaki’s Disease), MIS-A, and Long Covid seem to be products of the classical complement pathway (antibody mediated (exhausted IgG3 and the KKS. Not surprisingly Long Covid can frequently make a delayed appearance in MIS-C (10-30%)33. Asthma is the only reported pre-existing medical condition in pediatric Long Covid34.
In summary low levels of IFN-γ (due to exhausted CD4+and CD8+ T cells) with commensurately low levels of C1-INH result in activation of C1 and the CCP with crosstalk to the KKS. This creates an acquired angioedema clinically in those with increased Th1/Th2 and decreased IFN-γ, i.e., those that are vitamin D/Mg deficient and/or T cell exhausted => Long Covid.
1. What Population Does Long Covid Target?
If we approach the question from the perspective of an activated KKS, then bradykinin (BKN) should be the starting point.
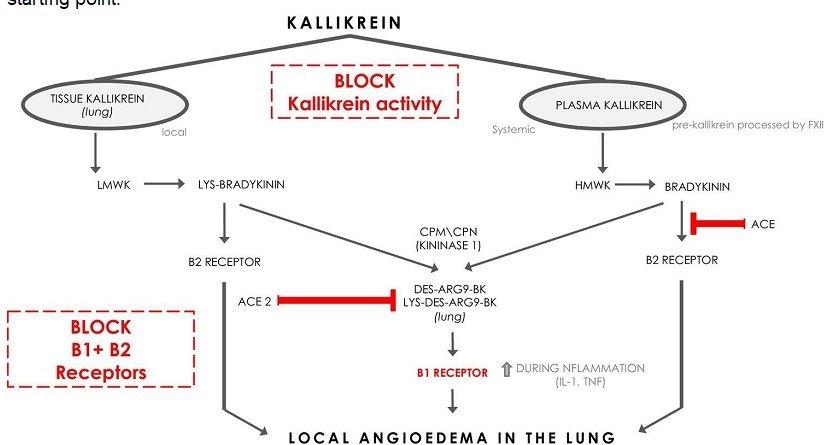
Figure 9. Pulmonary kallikrein is metabolized to des-Arg9-BK/lys-des-Arg9-BK, which will activate BKNR1s, if ACE2 has been sufficiently degraded35.
ACE is the main enzyme responsible for BKN degradation 36. The half-life of BKN is age independent. In men it is shorter than in women, because estrogen down-regulates ACE levels (vitamin D down-regulates estrogen)37. This means that Long Covid is more likely in women36. BKN operates via BKN2Rs and causes vascular leakage (Figure 9). If sufficient ACE2 receptors are degraded by SARS Cov2, then BKNIRs come into play (Figure 9). Loose endothelial intercellular junctions are associated with brain fog 38,39.
ACE levels for the ACE DD haplotype are up to 70% higher 40,41. The ACE DD haplotype is more common in African-Americans, which means that they are less likely to develop Long Covid. The evolutionary pressure of falciparum malaria favors the DD haplotype42 and provides protection against both Covid brain fog and childhood cerebral malaria 42,43. This explains the angioedema seen in many hypertensive African Americans on angiotensin converting enzyme inhibitors (ACEIs) 44. So both the ACE DD haplotype and COVID-19 enhance ACE and RAS activity but reduce BKN and KKS activity. The incidence of the ACE DD haplotype is greater in power sport athletes. On the other hand a high kinin ACE II haplotype has been linked to a common BKN2R polymorphism associated with enhanced endurance performance at an Olympic level, especially in triathlons(45).
According to a recent study, females appear to be more susceptible to Covid brain fog, 63% v 37% 43, the reverse of the Covid-19 gender breakdown. Age (median 43) and BMI (median 26) had no impact on cognitive function between those PCR+ and those PCR- (non hospitalized) long haulers, first seen for brain fog 5-6 months after onset of Covid-19 symptoms43. Covid brain fog seems to prefer Caucasians (88%) over African Americans (6%) 43. BKN can increase the permeability of the blood brain barrier 43 and the blood-gas barrier (lungs).
ACE/ACE2 provides a balance between angiotensin II and BKN with respect to thrombosis on the one hand and vascular permeability on the other. Too much ACE produces thrombotic microangiopathy (TMA). Too much BKN results in vascular leakage. Therefore, it looks like Long Covid prefers premenopausal females with the ACE II haplotype. Severe Covid-19 prefers males with the ACE DD haplotype (and those that live at altitude)46.
In summary the immunological differences between COVID-19 and long Covid are:
1. Severe Covid represents a problematic immune response, in which Th1>Th2 in responding to an invading pathogen, e.g., antibody independent. It involves mannose binding lectins (MBLs), MBL associated serine protease (MASP2), and the LCP 47 with activation of C2,4. Although it is strongly associated with vitamin D/magnesium deficiency, the KKS is uninvolved and not activated by the LCP (no crosstalk).
2. Long Covid and MIS-C/MIS-A represent a problematic immune response, in which Th1>Th2 in responding to Ag-Ab or immune complex, i.e., antibody dependent. This is a secondary response after mild to moderate primary exposure and recovery. It involves the classical complement pathway 47 , activates the KKS via C1 activation (crosstalk), and is strongly associated with vitamin D/magnesium deficiency.
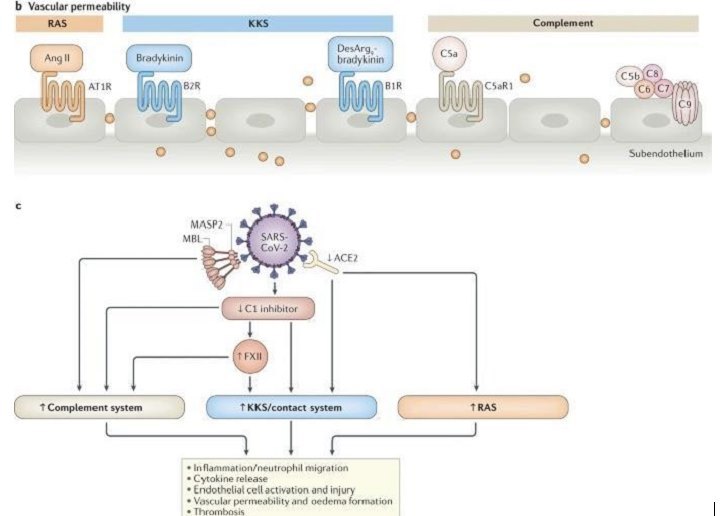
Figure 10. If IFN-γ is decreased and C1-INH with it, both Complement and the KKS are activated. If ACE2 is decreased, B1Rs are activated (Figure 9)48.
1. Is Long Covid One of the Faces of Magnesium Deficiency?
===Frequent symptoms of magnesium deficiency===
Fatigue
Migraines
Sleeplessness
Stress
Depression and anxiety
Hyperalgesia
Asthma
===Frequent symptoms of Long Covid===
Breathlessness
Fatigue
Brain fog
Hyperalgesia
Sleeplessness
Headaches
Asthma
1. What Other Faces Does Magnesium Deficiency Wear?
An alphabet soup of chronic inflammatory diseases seem to track back to a vitamin D/magnesium deficiency through decreased IFNγ expression 4996002-2"). IFN-γ signaling seems to be at the heart of many chronic inflammatory conditions, including EBV, CMV, Fibromyalgia, CFS/ME, and Long Covid50,51,52,53. Initially IFN-γ levels are high, but then the CD4+/CD8+ T cells producing the IFN-γ become exhausted with decreased IFN-γ. Although IFNγ plays a prominent role in autoimmune disease, its direction is controversial54. But what is clear is that vitamin D deficiency is involved in the pathogenesis of autoimmune disease55,56. Th17 cells and their IL17 cytokine, markers for Covid severity and autoimmune disease57, are unchallenged (see fig.8).
1. Conclusion
In summary, if 25(OH)D3 blood level is less than 50ng/ml, one is magnesium deficient. A vitamin D problem cannot be fixed in the face of a magnesium problem, e.g., vitamin D resistant magnesium deficient rickets. Modern osteoporosis seems to reflect a vitamin D resistant magnesium deficient entity. Surely this implies the existence of vitamin D resistant magnesium deficient immune dysfunction. The role of magnesium in human health cannot be underestimated and its role in immune function is complex. This discussion only scratches the surface. Genetic and epigenetic considerations that control Th1 and Th2 responses are in large measure a black box(58). Asthma paradoxically reduces risk for COVID-19 but increases it for MIS-C/MIS-A and Long Covid. But separation of allergic asthma from non-allergic asthma provides clarification. The eosinophils in the former and the smooth muscle bronchospasm in the latter shed light on this. Magnesium deficiency compromises interferon production, especially IFN-γ, and activates the CCP and KKS. IFN-γ links Long Covid with many other chronic inflammatory diseases. Indeed it links magnesium deficiency to dysfunctional immunomodulation. IFN-γ is also pleiotropic and can go both ways in autoimmune disease - pro-inflammatory in one circumstance yet anti-inflammatory in another, e.g., multiple sclerosis, Sjogren’s Syndrome59,60,61. IFN-γ can also promote tumor progression in one instance but regression in another62. Nonetheless, despite the pervasive ignorance, efforts toward achieving adequate vitamin D and magnesium seem prudent. Ironically the Frontline Covid Critical Care Alliance (FLCCC) makes no mention of magnesium in any of its protocols - general prevention, hospitalized treatment, long haul prevention. In fact only three items appear in all three protocols - vitamin D, vitamin C, and melatonin. Two of these require magnesium for their synthesis. Perhaps that is why they are on all three protocols. Almost everyone is short magnesium.
1. REFERENCES
1. Workinger, J. L., R. P. Doyle and J. Bortz (2018). "Challenges in the diagnosis of magnesium status." Nutrients 10(9). https://dx.doi.org/10.3390%2Fnu10091202
2. Cheung, M. M., R. DeLuccia, R. K. Ramadoss, A. Aljahdali, S. L. Volpe, P. A. Shewokis and D. Sukumar (2019). "Low dietary magnesium intake alters vitamin D-parathyroid hormone relationship in adults who are overweight or obese." Nutr Res 69: 82-93. https://doi.org/10.1016/j.nutres.2019.08.003
3. Ekwaru JP, Zwicker JD, Holick MF, Giovannucci E, Veugelers PJ (2014). “The Importance of Body Weight for the Dose Response Relationship of Oral Vitamin D Supplementation and Serum 25-Hydroxyvitamin D in Healthy Volunteers.” PLoS ONE 9(11): e111265. https://doi.org/10.1371/journal.pone.0111265
4. Ames, BN (2006). “The “triage theory”: micronutrient deficiencies cause insidious damage that accelerates age-associated chronic disease.” PNAS 103-17589-94 http://www.bruceames.org/Triage.pdf
5. Jeroen H. F. de Baaij, Joost G. J. Hoenderop, and René J. M. Bindels (2015). “Magnesium in Man: Implications for Health and Disease.” Physiological Reviews 2015 95:1, 1-46 https://doi.org/10.1152/physrev.00012.2014
6. Hosseini, A., H. Esmaeili Gouvarchin Ghaleh, H. Aghamollaei, M. Fasihi Ramandi, G. Alishiri, A. Shahriary, K. Hassanpour, M. Tat and G. Farnoosh (2021). "Evaluation of Th1 and Th2 mediated cellular and humoral immunity in patients with COVID-19 following the use of melatonin as an adjunctive treatment." Eur J Pharmacol 904: 174193. https://dx.doi.org/10.1016%2Fj.ejphar.2021.174193
7. Zou Z-G, Rios FJ, Montezano AC, Touyz RM (2019). “TRPM7, Magnesium, and Signaling.” International Journal of Molecular Sciences 20(8):1877. https://doi.org/10.3390/ijms20081877
8. Kim, M. H., S. Salloum, J. Y. Wang, L. P. Wong, J. Regan, K. Lefteri, Z. Manickas-Hill, C. Gao, J. Z. Li, R. I. Sadreyev, X. G. Yu, R. T. Chung, M. C. (2021). "Type I, II, and III interferon signatures correspond to coronavirus disease 2019 Severity." J Infect Dis 224(5): 777-782. https://doi.org/10.1093/infdis/jiab288
9. Chiang H-S and Liu HM (2019) “The Molecular Basis of Viral Inhibition of IRF- and STAT-Dependent Immune Responses.” Front. Immunol. 9:3086. https://doi.org/10.3389/fimmu.2018.03086
10. Goletti, D., L. Petrone, D. Manissero, A. Bertoletti, S. Rao, N. Ndunda, A. Sette, V. Nikolayevskyyet al. (2021) “The potential clinical utility of measuring severe acute respiratory syndrome coronavirus 2-specific T-cell responses.” Clin Microbiol Infect. 2021 Dec; 27(12): 1784–1789. https://doi.org/10.1016/j.cmi.2021.07.005
11. Ling, N., C. Meng-Li, F. Yu, Z. Hui, L. Jingyuan, Y. Fang, Y. Qing, Z. Gengzhen, et al. (2022) “Impaired Cellular Immunity to SARS-CoV-2 in Severe COVID-19 Patients.” Front.Immunol. 12:603563 https://dx.doi.org/10.3389%2Ffimmu.2021.603563
12. Cervia, C., Zurbuchen, Y., Taeschler, P. et al. (2022) “Immunoglobulin signature predicts risk of post-acute COVID-19 syndrome.” Nat Commun 13:446 https://doi.org/10.1038/s41467-021-27797-1
13. Luo H, Jia T, Chen J, Zeng S, Qiu Z, Wu S, Li X, Lei Y, et al (2021) “The Characterization of Disease Severity Associated IgG Subclasses Response in COVID-19 Patients.” Front. Immunol. 12:632814. https://dx.doi.org/10.3389%2Ffimmu.2021.632814
14. Wen, T.-H., Tsai, K.-W., Wu, Y.-J., Liao, M.-T., Lu, K.-C., Hu, W.-C. (2021) “The Framework for Human Host Immune Responses to Four Types of Parasitic Infections and Relevant Key JAK/STAT Signaling.” Int. J. Mol. Sci. 2021, 22, 13310. https://doi.org/10.3390/ijms222413310
15. Palacios-Gutiérreza, JJ, A Rodríguez-Guardadob, M Arias-Guillénc, R Alonso-Ariasd, S Palacios-Penedoe, J-M García-Garcíaf, et al (2022) “Clinical and epidemiological correlates of low IFN-gamma responses in mitogen tube of QuantiFERON assay in tuberculosis infection screening during the COVID-19 pandemic: A population-based marker of COVID-19 mortality?” Arch Bronconeumol. https://archbronconeumol.org/en-clinical-epidemiological-correlates-low-ifn-gamma-avance-S0300289622000825
16. Borekci, S., FG Karakas, S Sirekbasan, B Kubat, R Karaali, G (2021) “The Relationship between Pre-Pandemic Interferon Gamma Release Assay Test Results and COVID-19 Infection: Potential Prognostic Value of Indeterminate IFN-γ Release Assay Results.” Canadian Journal of Infectious Diseases and Medical Microbiology Volume 2021, Article ID 1989277, 9 pages https://dx.doi.org/10.1155%2F2021%2F1989277
17. Löbel, M., Mooslechner, A.A., Bauer, S. et al. (2015). “Polymorphism in COMT is associated with IgG3 subclass level and susceptibility to infection in patients with chronic fatigue syndrome.” J Transl Med 13, 264 https://doi.org/10.1186/s12967-015-0628-4
18. Hilgers A, Frank J. (1994) “Chronic fatigue syndrome: immune dysfunction, role of pathogens and toxic agents and neurological and cardiac changes.” Wien Med Wochenschr. 1994;144(16):399-406. http://www.ncbi.nlm.nih.gov/pubmed/7856214
19. Lotz M, Zuraw BL. (1987) “IFN-γ is a major regulator of C1-inhibitor synthesis by human blood monocytes.” J Immunol. 139(10):3382-3387. https://pubmed.ncbi.nlm.nih.gov/3119706/
20. Zuraw BL, Lotz M. (1990) “Regulation of the hepatic synthesis of C1 inhibitor by the hepatocyte stimulating factors interleukin 6 and interferon gamma.” J Biol Chem. 265(21):12664-12670. https://pubmed.ncbi.nlm.nih.gov/1695634/
21. Bossi, F., EI Peerschke, B Ghebrehiwet, F Tedesco (2011) “Cross-talk between the complement and the kinin system in vascular permeability.” Immunology Letters 140(1–2):7-13 https://doi.org/10.1016/j.imlet.2011.06.006
22. Hewison, M. (2010) “Vitamin D and the Immune System: New Perspectives on an Old Theme, Endocrinology and Metabolism.” Clinics of North America 39(2):365-379 https://doi.org/10.1016/j.ecl.2010.02.010
23. Maalmi H, Berraïes A, Tanguouru E, Ammar J, Abid H, Hamzaoui K, Hamzaoui A. (2012) “The impact of vitamin D deficiency on immune T cells in asthmatic children: a case-control study.” J Asthma Allergy. 5:11-19 http://dx.doi.org/10.2147/JAA.S29566
24. Agnes Hamzaoui, A. Berraïes, B. Hamdi, W. Kaabachi, J. Ammar, K. Hamzaoui (2014) “Vitamin D reduces the differentiation and expansion of Th17 cells in young asthmatic children.” Immunobiology 219(11):873-879 https://doi.org/10.1016/j.imbio.2014.07.009
25. Zhu, Z., K. Hasegawa, B. Ma, M. Fujiogi, CA Camargo Jr., L. Liang (2020) “Association of asthma and its genetic predisposition with the risk of severe COVID-19.” Journal of Allergy and Clinical Immunology 146(2):327–329. https://doi.org/10.1016/j.jaci.2020.06.001
26. Ferastraoaru D, Hudes G, Jerschow E, Jariwala S, Karagic M, de Vos G, Rosenstreich D, Ramesh M. (2021) “Eosinophilia in Asthma Patients Is Protective Against Severe COVID-19 Illness.” J Allergy Clin Immunol Pract. 9(3):1152-1162. https://doi.org/10.1016/j.jaip.2020.12.045
27. Shaikh, MN, Malapati, BR, Gokani, R, Patel, B, Chatriwala, M (2016) “Serum Magnesium and Vitamin D Levels as Indicators of Asthma Severity.” Pulm Med. 2016:1643717. https://dx.doi.org/10.1155%2F2016%2F1643717
28. Zhong-Jie, H., X. Jia, Y. Ji-Ming, L. Li, H. Wei, Z. Li-Li, Z. Zhen, et al. (2020) “Lower Circulating Interferon-Gamma Is a Risk Factor for Lung Fibrosis in COVID-19 Patients.” Frontiers in Immunology 11:585647. https://dx.doi.org/10.3389%2Ffimmu.2020.585647
29. Abdel-Hamed, E.F., Ibrahim, M.N., Mostafa, N.E. et al. (2021) “Role of interferon gamma in SARS-CoV2 positive patients with parasitic infections.” Gut Pathog 13:29 https://doi.org/10.1186/s13099-021-00427-3
30. Sapkota, H.R., Nune, A. (2022) “Long COVID from rheumatology perspective — a narrative review.” Clin Rheumatol 41:337–348 https://dx.doi.org/10.1007%2Fs10067-021-06001-1
31. Nabi-Afjadi, M., Karami, H., Goudarzi, K. et al. (2021) “The effect of vitamin D, magnesium and zinc supplements on interferon signaling pathways and their relationship to control SARS-CoV-2 infection.” Clin Mol Allergy 19:21. https://doi.org/10.1186/s12948-021-00161-w
32. Porritt, RA, A Binek, L Paschold, MN Rivas, A McArdle, LM Yonker, et al. (2021) “The autoimmune signature of hyperinflammatory multisystem inflammatory syndrome in children.” J Clin Invest. 131(20):e151520. https://doi.org/10.1172/JCI151520
33. Rubin L, Subramony A, Kothare S. (2022) “Patients Diagnosed with Multisystem Inflammatory Syndrome in Children Have Persistent Neurologic, Sleep, and Psychiatric Symptoms After Hospitalization” J Child Neurol. 24:8830738221075924. https://doi.org/10.1177/08830738221075924
34. Zimmermann, P., Pittet, LF, Curtis, N., (2021) “How Common is Long COVID in Children and Adolescents?” The Pediatric Infectious Disease Journal: December 40(12):e482-e487 https://dx.doi.org/10.1097%2FINF.0000000000003328
35. van de Veerdonk, FL, Netea, MG, van Deuren, M, van der Meer, JWM, de Mast, Q, et al. (2020) “KKS blockade in patients with COVID-19 to prevent acute respiratory distress syndrome.” eLife 2020;9:e57555. https://doi.org/10.7554/eLife.57555
36. Cyr, M., Y. Lepage, C. Blais, Jr., N. Gervais, M. Cugno, J-L. Rouleau, A.Adam (2001) “Bradykinin and des-Arg9-bradykinin metabolic pathways and kinetics of activation of human plasma.” American Journal of Physiology-Heart and Circulatory Physiology 281(1):H275-H283 https://doi.org/10.1152/ajpheart.2001.281.1.H275
37. Gallagher, P. E., Li, P., Lenhart, J. R., Chappell, M. C., K. B. Brosnihan (1999) “Estrogen Regulation of Angiotensin-Converting Enzyme mRNA.” American Heart Association 33(1):323-328. https://doi.org/10.1161/01.hyp.33.1.323
38. Schwaninger, M., S. Sallmann, N. Petersen, A. Schneider, S. Prinz, T.A. Libermann, M. Spranger (1999) “Bradykinin Induces Interleukin-6 Expression in Astrocytes Through Activation of Nuclear Factor-κB.” J. Neurochem. 73,1461–1466 https://doi.org/10.1046/j.1471-4159.1999.0731461.x
39. Bas, M., TK Hoffmann, B Tiemann, VTV Dao, C Bantis, V Balz, H-J Schultz-Coulon, T Stark, et al. (2010) “Potential genetic risk factors in angiotensin-converting enzyme-inhibitor-induced angio-oedema.” Br J Clin Pharmacol 69(2):179–186 https://doi.org/10.1111/j.1365-2125.2009.03567.x
40. Biller, H., Zissel, G., Ruprecht, B., Nauck, M., Busse Grawitz, A., Müller-Quernheim, J. (2006) “Genotype-corrected reference values for serum angiotensin-converting enzyme.” European Respiratory Journal 28:1085-1091. https://doi.org/10.1183/09031936.00050106
41. Rigat, B., Corvol, P., Soubrier, F. (1990) “An insertion/deletion polymorphism in the angiotensin I converting enzyme gene accounting for half the variance of serum enzyme levels.” J Clin Invest. 86(4):1343-1346. https://doi.org/10.1172/JCI114844
42. Gallego-Delgado, J., Walther, T., Rodriguez, A. (2016) “The High Blood Pressure-Malaria Protection Hypothesis.” Circulation Research. 119:1071–1075 https://doi.org/10.1161/CIRCRESAHA.116.309602
43. Graham, EL, Clark, JR, Orban, ZS, Lim, PH, Szymanski, AL, et al, (2021) “Persistent neurologic symptoms and cognitive dysfunction in non hospitalized Covid19 long haulers.” Annals of Clinical and TranslationalNeurology2021; 8(5): 1073–1085. https://doi.org/10.1002/acn3.51350
44. Teuwen, LA., Geldhof, V., Pasut, A. et al. (2020) “COVID-19: the vasculature unleashed.” Nat Rev Immunol 20, 389–391 https://doi.org/10.1038/s41577-020-0343-0
45. Puthucheary, Z., Skipworth, J.R.A., Rawal, J. et al. (2011) “The ACE Gene and Human Performance.” Sports Med 41:433–448 https://doi.org/10.2165/11588720-000000000-00000
46. Chambers, P.W. (2022) “COVID-19: From Cough to Coffin.” Open Access Library Journal, 9:1-24. https://doi.org/10.4236/oalib.1108300
47. Java, A., AJ Apicelli, K Liszewski, A Coler-Reilly, JP Atkinson, AHJ Kim, HS Kulkarni (2020) “The complement system in COVID-19: friend and foe?” JCI Insight. 5(15):e140711 https://doi.org/10.1172/jci.insight.140711
48. Bekassy, Z., Lopatko Fagerström, I., Bader, M. et al. (2021) “Crosstalk between the renin–angiotensin, complement and kallikrein–kinin systems in inflammation.” Nat Rev Immunol. https://doi.org/10.1038/s41577-021-00634-8
49. Schoenborn, J.R., CB Wilson. (2007) “Regulation of Interferon‐γ During Innate and Adaptive Immune Responses.” Advances in Immunology 96:41-101 https://doi.org/10.1016/s0065-2776(07)96002-296002-2)
50. Rosa Bellmann-Weiler, Katharina Schroecksnadel, Claudia Holzer, Clara Larcher, Dietmar Fuchs, Guenter Weiss (2008) “IFN-gamma mediated pathways in patients with fatigue and chronic active Epstein Barr virus-infection.” Journal of Affective Disorders 108(1-2):171-176 https://doi.org/10.1016/j.jad.2007.09.005
51. Sinclair, E., D Black, CL Epling, A Carvidi, SZ Josefowicz, BM Bredt, MAVJacobson (2004) “CMV Antigen-Specific CD4+ and CD8+ T Cell IFNγ Expression and Proliferation Responses in Healthy CMV-Seropositive Individuals.” Viral Immunology 17(3):445-454 https://dx.doi.org/10.1089%2F0882824041857049
52. Björkander, S., M Ernberg, I Bileviciute-Ljungar (2022) “Reduced immune system responsiveness in fibromyalgia - A pilot study.” Clinical Immunology Communications 2:46-53 https://doi.org/10.1016/j.clicom.2022.02.003
53. Deumer, U.-S., Varesi, A., Floris, V., Savioli, G., Mantovani, E., López-Carrasco, P., Rosati, G.M., Prasad, S., Ricevuti, G. (2021) “Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): An Overview.” J. Clin. Med. 10:4786. https://dx.doi.org/10.3390%2Fjcm10204786
54. Arellano G, Ottum PA, Reyes LI, Burgos PI and Naves R (2015) “Stage-specific role of interferon-gamma in experimental autoimmune encephalomyelitis and multiple sclerosis.” Front. Immunol. 6:492. https://doi.org/10.3389/fimmu.2015.00492
55. Passwater, M. (2022) “It’s Official: Vitamin D Reduces the Incidence of Autoimmunity.” Orthmolecular.org v18(9) http://orthomolecular.org/resources/omns/v18n09.shtml
56. Zhao R, Zhang W, Ma C, Zhao Y, Xiong R, Wang H, Chen W, Zheng SG (2021) “Immunomodulatory Function of Vitamin D and Its Role in Autoimmune Thyroid Disease.” Front. Immunol. 12:574967. https://dx.doi.org/10.3389%2Ffimmu.2021.574967
57. Pourgholaminejad A., S.Pahlavanneshan, M. Basiri (2022) “COVID-19 immunopathology with emphasis on Th17 response and cell-based immunomodulation therapy: Potential targets and challenges.” Scand J Immunol. 95:e13131. https://doi.org/10.1111/sji.13131
58. Sanders, VM (2006) “Epigenetic regulation of Th1 and Th2 cell development.” Brain, Behavior, and Immunity 20(4):317-324 https://doi.org/10.1016/j.bbi.2005.08.005
59. Zhang J. (2007) “Paradoxical roles of interferon-γ in autoimmune disease.” Expert Review of Clinical Immunology, 3(1):35-38 https://doi.org/10.1586/1744666x.3.1.35
60. Kelchtermans, H., A Billiau, P Matthys (2008) “How interferon-γ keeps autoimmune diseases in check.” Trends in Immunology 29(10):479-486 https://doi.org/10.1016/j.it.2008.07.002
61. Ottum P A., G Arellano, LI Reyes, M Iruretagoyena, Rodrigo N (2015) “Opposing Roles of Interferon-Gamma on Cells of the Central Nervous System in Autoimmune Neuroinflammation.” Frontiers in Immunology 6:539. https://doi.org/10.3389/fimmu.2015.00539
62. Mandai, M., Hamanishi, J., Abiko, K., Matsumura, N., Baba, T., & Konishi, I. (2016). “Dual Faces of IFNγ in Cancer Progression: A Role of PD-L1 Induction in the Determination of Pro- and Antitumor Immunity.” Clinical cancer research (an official journal of the American Association for Cancer Research) 22(10):2329–2334. https://doi.org/10.1158/1078-0432.CCR-16-0224
1. VitaminDWiki - <a href="/pages/magnesium-and-vitamin-d/">Magnesium and Vitamin D</a> contains
{include}
1. VitaminDWiki - <!-- {SQL(db=>vitamind)}SELECT COUNT(*) FROM (SELECT catObjectId, COUNT(*) FROM `tiki_category_objects` WHERE categId = 155 OR categId = 54 GROUP BY catObjectId HAVING COUNT(*) > 1) AS categs INNER JOIN tiki_objects ON tiki_objects.objectId = categs.catObjectId WHERE type = "wiki page"{SQL} --> studies in both categories Virus and Magnesium
This list is automatically updated
{category}
{include}
1. There have been <span style="color:#00F;"><!-- {SQL(db=>vitamind)}SELECT hits FROM tiki_pages WHERE page_id=13452{SQL} --></span> visits to this page