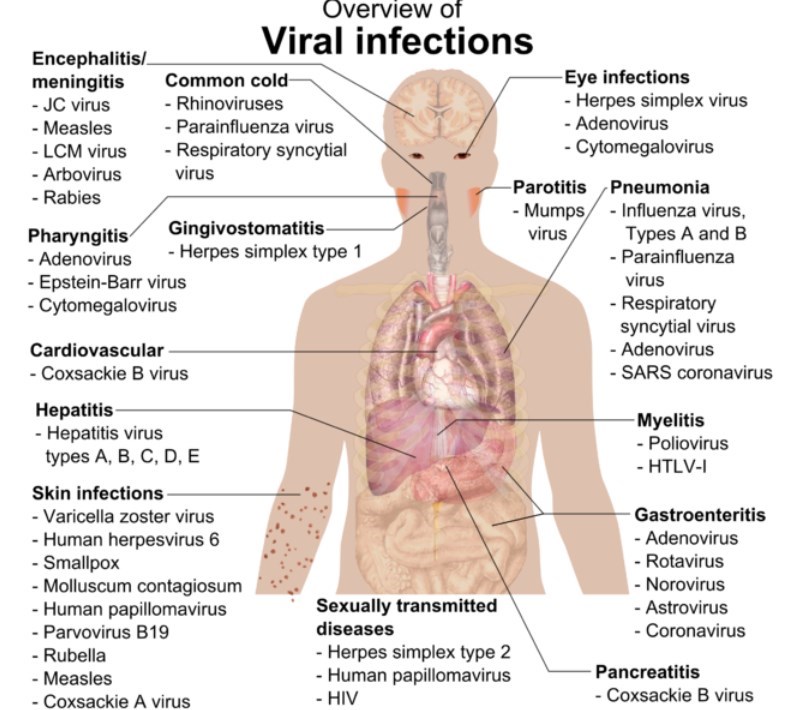
Vitamin D can inhibit enveloped virus (e.g. Corona, Herpes, Bird Flu, Epstein, Hepatitis, RSV, etc.)
Examples of enveloped virus Coronavirus, Herpes simplex (cold sore), Shingles, Epstein-Barr, Hepatitis A & B & C, HMPV, Zika, HIV, Dengue, Yellow fever, Measles, Mumps, smallpox, Monkeypox, Ebola, rabies, influenza, West Nile, Valley Fever, Croup, Lasa Fever, Hemorrhagic Fever, H5N1 bird flu, Parkinson's Disease, African Swine Flu
Vitamin D and the anti-viral state - March 2011
J Clin Virol. 2011 March; 50(3): 194–200.
Jeremy A. Beard,b Allison Bearden,a,b and Rob Striker a,b, rtstriker@wisc.edu (608) 263-2994
a W. S. Middleton Memorial Veterans Administration Hospital, 53705, USA http://www.madison.va.gov/
b University of Wisconsin-Madison, Department of Medicine, 53706, USA https://www.medicine.wisc.edu/
Photo of Rob Stiker

Vitamin D has long been recognized as essential to the skeletal system. Newer evidence suggests that it also plays a major role regulating the immune system, perhaps including immune responses to viral infection. Interventional and observational epidemiological studies provide evidence that vitamin D deficiency may confer increased risk of influenza and respiratory tract infection. Vitamin D deficiency is also prevalent among patients with HIV infection. Cell culture experiments support the thesis that vitamin D has direct anti-viral effects particularly against enveloped viruses . Though vitamin D’s anti-viral mechanism has not been fully established, it may be linked to vitamin D’s ability to up-regulate the anti-microbial peptides LL-37 and human beta defensin 2 . Additional studies are necessary to fully elucidate the efficacy and mechanism of vitamin D as an anti-viral agent.
1. Introduction and general physiology
Vitamin D is known for its traditional role in bone mineralization and calcium homeostasis. It is an essential part of the human diet. The body can handle doses as high as 10,000 IU (250 μg) per day for several months.1 Mounting evidence suggests that it plays a major role in mediating the immune system’s response to infection.2 Therefore, vitamin D represents a potentially useful intervention for combating viral infection. Further study may aid in understanding the role of vitamin D in viral pathogenesis. The vast literature on vitamin D includes reviews on many topics including its effects on innate immunity,3 cardiovascular health,4 and cancer.5 This review focuses on vitamin D’s putative role in establishing a preventative and therapeutic anti-viral state.
Vitamin D exists in several forms including 25-hydroxyvitamin D [25(OH)D], the primary circulating form, and 1,25-dihydroxyvitamin D [1,25(OH)2D], the active form.1 Vitamin D is obtained by skin exposure to sunlight (thereby converting 7-dehydrocholesterol to cholecalciferol, vitamin D3), from foods, or through supplements. It can be ingested in the form of vitamin D3 or vitamin D2 (ergocalciferol). Vitamin D2 is derived from irradiation of the fungal steroid ergosterol.6 After digestion, vitamin D is processed by 25-hydroxylases present in the liver and other tissues to generate 25-hydroxyvitamin D [25(OH)D].7–9 Subsequently, 25-hydroxyvitamin D is converted to 1,25-dihydroxyvitamin D by the enzyme 25-hydroxyvitamin D-1-α-hydroxylase, CYP27B1.10,11
Serum 25(OH)D correlates with overall vitamin D stores and is the most commonly used biomarker for assessing vitamin D deficiency.10–14 Deficiency is often defined by circulating 25(OH)D levels below 20 ng/ml (50 nmol/l).11,15,16 but 30 ng/ml (75 nmol/l)17,18 and even 40 ng/ml (100 nmol/l) are sometimes advocated for specific patients.19–21
1,25-Dihydroxvitamin D [1,25(OH)2D] is primarily generated in the kidneys by a 1-α-hydroxylase, CYP27B1.10,11 CYP27B1 is also present in a variety of extra-renal tissues including immune cells, and unlike the renal form of the enzyme, is not regulated by calcium metabolism signaling.22–24 CYP27B1 in keratinocytes is up-regulated in response to injury, and toll-like receptor (TLR) activation by microbial products.25,26 In addition, activated macrophages, dendritic cells, T lymphocytes, and B lymphocytes express CYP27B1.27–30 Catabolism of vitamin D is accomplished by 24-hydroxylases including CYP24A1. A negative feedback loop exists as catabolic enzymes are induced by 1,25(OH)2D.10
Given that 1,25(OH)2D is the active form of vitamin D, it is tempting to use this for diagnosis and monitoring of vitamin D status. Such an approach can be problematic. Due to its increased biological half-life and other factors, 25(OH)D is normally present in higher concentrations than its active metabolite. However, vitamin D deficiency results in increased parathyroid hormone, inducing renal hydroxylation of 25(OH)D via renal CYP27B1.14 This additional regulation of vitamin D by calcium and parathyroid hormone can result in normal or elevated 1,25(OH)2D levels despite systemic vitamin D deficiency.14,22,23
2. Vitamin D molecular mechanisms and immune modulation
The effects of 1,25(OH)2D are mediated by it binding to the vitamin D receptor (VDR). VDR is a nuclear receptor and once it binds its ligand, VDR dimerizes with an isoform of the retinoid X receptor. These VDR-RXR heterodimers bind to vitamin D response elements present on target genes.31–33 In addition to transcriptional activation, the heterodimers can displace the nuclear factors of activated T cells resulting in repression of cytokine related genes.34
1,25(OH)2D suppresses Th-1 cell proliferation leading to lowered production of interferon gamma and interleukin-2.27,35,36 Lower levels of circulating cytokines leads to less antigen presentation by dendritic cells, in addition to less T lymphocyte recruitment and proliferation.36 Expression of Th-2 associated cytokines, including interleukin-4 are increased by 1,25(OH).37 Overall, vitamin D polarizes the adaptive immune system away from Th-1 and toward Th-2 responses.
Vitamin D also plays a role in innate immune response modulation. The toll-like receptors (TLRs) in macrophages, polymorphonuclear cells, monocytes, and epithelial cells are central to the innate immune response.38,39 TLRs recognize pathogen associated molecular patterns associated with infectious agents.39 For example, TLR2 recognizes the lipopolysaccharides of bacteria. TLRs have also been shown to recognize viral proteins and nucleic acids.40 Upon recognition, activated TLRs release cytokines that induce expression of antimicrobial peptides and reactive oxygen species.
Several TLRs both affect and are affected by VDR stimulation. Expression of CD-14, the co-receptor for TLR4, is induced by 1,25(OH)2D in monocytes and epidermal keratinocytes.26,41 Stimulation of TLR2 in macrophages by anti-microbial peptides leads to increased local expression of CYP27B1, resulting in the conversion of vitamin D to its active form.25 Some anti-microbial peptides associated with TLRs have demonstrated anti-viral effects, and their expression is affected by vitamin D levels.42 Human beta defensin 2 is modestly up-regulated by 1,25(OH)2D and may contribute to anti-viral effects as a chemoattractant for neutrophils and monocytes.38,43 Conversely, in monocytes activation by 1,25(OH)2D alone is insufficient for induction of gene expression.44 Human cathelicidin is an antimicrobial peptide induced by TLR1/2 activation. Cathelicidin is strongly up-regulated by 1,25(OH)2D due to the its VDR response element.44–46
Cathelicidins are a family of proteins with a C-terminal cationic anti-microbial domain activated by cleavage from the N-terminal cathelin domain.46 In humans, the active antimicrobial cathelicidin peptide LL-37 is cleaved from the propeptide, hCAP18.47 Although the majority of cathelicidin is stored in neutrophil granules for release at sites of infection, several other types of immune cells including monocytes, NK cells, and B cells express hCAP18.48 It is secreted into the blood and by the epithelia of the conjuctiva, cornea, respiratory tract, digestive tract, epithelial tract, intestines, urinary tract, and skin.49–52 At the cellular level, expression of CYP27B1 in macrophages and keratinocytes induces cathelicidin expression.14,25 If there is no 25(OH)D, VDR, or CYP27B1 present, the ability of these cell types to induce cathelicidin is significantly impaired.25,26
In addition to anti-bacterial effects including membrane disruption25,51–53, cathelicidin in the peptide form LL-37, has demonstrated anti-viral effects including inhibition of herpes simplex virus type one (HSV-1), vaccinia virus replication, retroviral replication, and replication of some adenovirus serotypes at certain peptide concentrations.50,54–56
3. Evidence for role of vitamin D in viral respiratory infections
Recent work highlights vitamin D’s potential role in fighting viral respiratory infections. Lung epithelial cells express high basal levels of CYP27B1 and low levels of CYP24A1, favoring conversion of vitamin D to its active form.57 When treated with vitamin D, these cells increase the levels of the TLR co-receptor CD-14 and cathelicidin.57 In airway epithelial cells, treatment with vitamin D induces IkBα, an NF-kB inhibitor resulting in a decrease of viral induction of inflammatory genes.58
Studies have identified possible links between vitamin D and respiratory infections by examining VDR polymorphisms. Single nucleotide polymorphisms in VDR and related genes are associated with severe outcomes in respiratory syncytial virus (RSV) related bronchiolitis and acute lower respiratory tract infection (RTI) likely due to VDR association with innate immunity.59,60
Controlled trials examining the effect of vitamin D supplementation on reducing RTIs have had mixed results. A 1994 study done in India showed a reduction in respiratory infections of 27 children treated for six weeks with vitamin D.61 These children had a previous history of RTIs and vitamin D deficiency. A British study of 1740 elderly patients administered 800 IU over a two year period showed no significant reduction in infections compared to controls.62 A New York study involving a mostly Caucasian population showed daily administration of 2000 IU of vitamin D3 had no significant effect on decreasing the incidence or severity of respiratory tract infections during winter.63
In the New York study, the serum mean of 25(OH)D was above deficiency levels. In addition, the subjects did not begin vitamin D supplementation prior to the wintertime. As the authors note, given that it can take up to three months for 25(OH)D levels to reach a steady state with supplementation, this may have influenced the study result.64,65 These results suggest the effect is most pronounced or only present in vitamin D deficient patients. Differences between the trials might result from patient underreporting of RTIs. Many relied on patient questionnaires and not clinical diagnosis. The results of the controlled trials are summarized in Table 1.
Table 1
Controlled studies of vitamin D based treatment for prevention of respiratory tract infections and influenza.
Observational studies evaluating the relationship between serum 25(OH)D concentrations and respiratory infections also have had mixed results. A Finnish study found an association between serum 25(OH)D concentrations less than 16 ng/ml (40 nmol/l) and an increased incidence of acute respiratory tract infections.66 A two month study of Bangladeshi children found a significant correlation between increased numbers of RTIs and significantly lower mean levels of 25(OH)D of 11.7 ng/ml (29.1 nmol/l) versus 15.7 ng/ml for controls (39.1 nmol/l).67 Similar results were found in studies of Indian and Turkish children.68,69 Two Canadian studies of children showed no significant difference in mean 25(OH)D levels between RTI patients and controls.70,71 A retrospective analysis of the Third National Health and Nutrition Examination Survey of 18,883 patients showed that 25(OH)D levels less than 30 ng/ml (75 nmol/l) were associated with an increased risk of upper respiratory tract infection.72 Patients with levels less than 10 ng/ml had a 55% risk of infection when compared to controls. Again, this suggests that if a patient is not vitamin D deficient, there is limited anti-viral benefit gained from supplementation. The results of the observational studies are summarized in Table 2.
Table 2
Observational studies correlating vitamin D deficiencies with respiratory tract infection and influenza.
While researchers have suggested a link between seasonal variation in vitamin D levels and influenza,73 a Japanese supplementation trial during the winter and early spring showed only a mild reduction in influenza A infections in children taking vitamin D3 supplements. However, the study used only outpatients and did not measure serum concentrations of 25(OH)D or serum antibody concentrations to influenza A. It is possible that the milder forms of disease and the extreme forms requiring hospitalization were not recorded.74
The Japanese study’s homogeneous population means the correlation between mild reduction of influenza A and vitamin D supplementation cannot easily be generalized as skin pigmentation impacts vitamin D production.73,75 Therefore, darker skin individuals may gain more benefit from supplementation. For example, in a three-year study of post-menopausal African-American women receiving vitamin D supplementation, researchers found a reduction in reported cold and influenza.76
4. Evidence for vitamin D influence on HIV infection
Observational studies have reported lower levels of vitamin D in HIV populations. In a German study, 25(OH)D levels of less than 20 ng/ml (50 nmol/l) were found in 47.6% of the subjects with AIDS.77 Another study of 50 women with HIV found significantly lower 1,25(OH)2D levels in the patients compared to healthy female controls.78 In a study of HIV-infected adults from the United States, serum levels of 25(OH)D were below normal values in only 17% of subjects and the 1,25(OH)2D serum levels were low in 11% of subjects though the differences were of only borderline statistical significance.79 A Norwegian study of 53 patients also found significantly lower 1,25(OH)2D serum levels than controls.80 Interestingly, in this Norwegian study the serum concentrations of 25(OH)D were not significantly lower than that of the controls. Even when patients being treated with drugs known to inhibit CYP27B1 were excluded, the deficiency in the cohort persisted.80 This suggests a novel mechanism.
Although the studies (summarized in Table 3) show an association between lower vitamin D levels and HIV infection, they do not clarify the nature of that relationship. As the active form of vitamin D, 1,25(OH)2D is typically more reduced than 25(OH)D, it is unlikely this is solely because of diet and sunlight exposure. However, in some patients, pre-infection vitamin D levels were low because of such factors.81 One possible mechanism put forth to explain the vitamin D deficiencies is that over activation of TNF-α in HIV patients might lead to a blocking of the stimulatory effect of parathyroid hormone on renal 1-α-hydroxylase.82
Table 3
Observational studies examining vitamin D deficiencies and HIV infection.
As some anti-retroviral drugs, including protease inhibitions, have been shown to interfere with vitamin metabolism in vitro,83 several studies have examined the potential impact of HAART regimens on vitamin D levels in HIV patients. These studies have found associations between low vitamin D levels and the use of non-nucleoside reverse transcriptase inhibitors and protease inhibitors.84–88 However, at least one study supported the idea that though protease inhibitors are associated with lower 1,25(OH)2D levels, they were not the cause of vitamin D deficiency.89
Due to the different effects of anti-virals on vitamin D metabolism and cohort effects, the clinical implication of low vitamin D levels in HIV patients is not clear. Observational studies of humans have found positive correlations between vitamin D and CD4+ levels.78,80,82,90,91 At least one study has found a correlation between higher vitamin D levels and increased survival times of HIV-infected patients.80 However, a 2004 study by Madeddu et al. of 152 adult patients on HAART, found no correlation between vitamin D and CD4+ T cell levels.89 A 2001 study of 19 perinatally infected female children also found no correlation between vitamin D and CD4+ T cell levels.92
5. Evidence for vitamin D influence on Epstein Barr virus
Studies have suggested a link between multiple sclerosis (MS) and Epstein-Barr virus (EBV), thus vitamin D levels may play a role in the development of MS.93,94 This topic was reviewed by Trygve Holmoy in Medical Hypotheses.95 Holmoy notes that MS risk is associated with low vitamin D status and EBV infection. He proposes that vitamin D modulates the immune response to EBV and suppresses activation of auto-reactive T cells that may contribute to MS pathology.
6. Other evidence for vitamin D influence on enveloped viruses
Although few studies have examined the effects of vitamin D on Hepatitis B infection, a study of 2015 Gambian tuberculosis patients identified a silent T to C base change polymorphism in codon 352 of the VDR that was correlated with significantly lowered rates of persistent Hepatitis B infection and tuberculosis, but not malaria.96 This polymorphism affects vitamin D levels, VDR mRNA stability, and VDR mRNA levels.97–100 The anti-Hepatitis B response in these patients may be cathelicidin mediated, much like the cathelicidin-mediated anti-tuberculosis response recently described.25
In a Vietnamese dengue study, the same polymorphism was associated with resistance to severe dengue.101 At least one study has demonstrated administration of oral vitamin D3 reduces the severity and length of dengue fever, but the small sample size (n = 5) makes it difficult to draw any robust conclusions.102
A study conducted by Bitetto et al. showed that in immune competent patients 25(OH)D levels of less than 10 ng/ml (25 nmol/l) are significantly associated with poorer response to the standard antiviral hepatitis C therapy of ribavirin and pegylated interferon.103 An earlier study by Petta et al. revealed an association between lower vitamin D levels and failure to clear virus during treatment in chronic hepatitis C patients.104 The same study showed that low vitamin D levels were also associated with liver fibrosis. Therefore, this makes it difficult to determine whether lower vitamin D levels correlate with lower viral clearance or are just a consequence of damaged livers.
7. Potential mechanisms of anti-viral effects
The anti-viral effects of vitamin D could be explained by cathelicidin (in the form of LL-37), human beta defensin 2, and perhaps through the release of reactive oxygen species. A recent study showed hepatitis C replicon replication reduction in human hepatoma cells may be mediated by vitamin D induced oxidative stress.105 Given vitamin D’s pleiotropic effects, other mechanisms are possible.
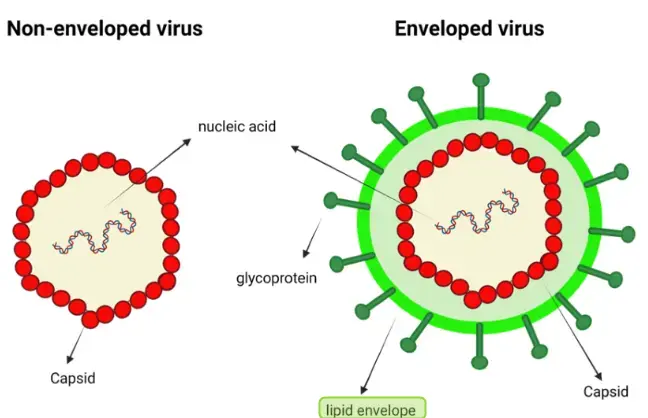
LL-37’s anti-bacterial effect is linked to its ability to disrupt bacterial membranes through electrostatic interactions.53 Similar interactions may occur with the lipid envelopes of viruses. LL-37 may also block viral entry in a similar manner to what is seen with other anti-microbial peptides.106 The epidemiologic evidence describing a positive vitamin D related immune effect includes many studies which feature enveloped viruses. This supports the notion that LL-37’s anti-viral effects may be partially mediated by envelope disruption.
8. Experimental investigation of the anti-viral effects of vitamin D
Vitamin D induction of antimicrobial peptides may have antiviral effects.43 Direct incubation of LL-37 with HSV-1 showed a significant dose dependent reduction of HSV-1 titer when compared to controls.50 The same researchers also demonstrated a less pronounced but still significant reduction of adenovirus serotype Ad19 titer when exposed to higher LL-37 concentrations, but no significant titer reduction of the other adenovirus serotypes tested (Ad8, Ad5 and Ad3). Human papilloma virus appears sensitive to LL-37 inactivation or entry inhibition at physiological concentrations of LL-37, but some retroviruses are also sensitive to LL-37 mediated titer reduction at concentrations that may not be physiologically relevant.54,56,107 Methodological differences may affect these comparisons.
While mouse models do not perfectly parallel vitamin D’s effects on human anti-microbial peptides, they provide evidence that anti-microbial peptides may have an effect against viruses. Studies involving incubation of LL-37 or CRAMP, a murine homolog, with vaccinia showed an almost complete inhibition, as measured by titer, of the virus at a concentration of 50 micromolar.54 The vaccinia experiments also showed that in CRAMP knockout mice inoculated with skin pricks of vaccinia, four of six knockout animals developed vaccinia pox compared to only one of fifteen controls. While beta defensin 2 was unable to reduce vaccinia titer,54 it is capable of inhibiting RSV in lung epithelial cell culture. Transmission electron microscopy showed the degradation of the RSV membrane after incubation with human defensin beta 2.108 Mice infected with RSV expressed increased levels of murine beta defensin 4, a murine analog of human beta defensin 2, suggesting the cell culture results have in vivo relevance.
Go to:
9. Conclusions
These results support the hypothesis that vitamin D induced LL-37, and to a lesser extent human beta defensin 2, may play a major role in the inhibition of viruses. However, these experiments do not completely model the complex effects of vitamin D and may not accurately represent its systemic influence. Further experiments are necessary to fully elucidate the mechanisms of vitamin D induced peptides and vitamin D itself.
References (missing the first 16) all references are in the PDF
Chapuy MC, Preziosis P, Maamer M, Arnaud S, Galan P, Hercberg Meunier PJ. Prevalance of vitamin D insufficiency in an adult normal population. Osteoporos Int. 1997; 7:439-44. [PubMed: 9425501]
Thoma-Uszynsk SS, Lloyd Jones M, Thadhani RI, Shaw AC, Deraska DJ, Kitch BT, Vamvakas EC, Dick IM, Prince RL, Finkelstein JS. Hypovitaminosis D in medical inpatients. N Engl J Med. 1998; 338:777-83. [PubMed: 9504937]
Binkley N, Krueger D, Gemar D, Drezner MK. Correlation among 25-hydroxy-vitamin D assays. J Clin Endocrinol Metab. 2008; 93(5):1804-8. [PubMed: 18319318]
McKenna MJ, Freaney R. Secondary hyperparathyroidism in the elderly: means to defining hypovitaminosis D. Osteoporos Int. 1998; 8(Suppl 2):S3-6. [PubMed: 10197175]
Holick MF. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers, and cardiovascular disease. Am J Clin Nutr. 2004; 80(Suppl 6):1678S-88S. [PubMed: 15585788]
Batchelor AJ, Compston J. Reduced plasma half-life of radio-labeled 25-hydroxyvitamin D, in subjects receiving a high-fiber diet. Br J Nutr. 1983; 49:213-5. [PubMed: 6299329]
Holick F. Vitamin D: importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am J Clin Nutr. 2004; 79(3):362-71. [PubMed: 14985208]
White JH, Vitamin D, Signaling. Infectious diseases, and regulation of innate immunity. Infect Immun. 2008; 76(9):3837-43. [PubMed: 18505808]
Liu PT, Stenger S, Li H, Wenzel L, Tan BH, Krutzik SR, et al. Toll-like receptor triggering of a vitamin D-mediated anti-microbial response. Science. 2006; 311(5768):1770-3. [PubMed: 16497887]
Schauber J, Dorschner RA, Coda AB, Buchau AS, Liu PT, Kiken D, et al. Injury enhances TLR2 function and anti-microbial peptide expression through a vitamin D-dependent mechanism. J Clin Invest. 2007; 117(3):803-11. [PubMed: 17290304]
Chen S, Sims GP, Chen XX, Gu YY, Chen S, Lipsky PE. Modulatory effects of 1,25- dihydroxyvitamin D3 on human B cell differentiation. J Immunol. 2007; 179:1634-47. [PubMed: 17641030]
Sigmunsdottir H, Pan J, Debes GF, Alt C, Habtezion A, Soler D, et al. DCs metabolize sunlight inducted vitamin D3 to ‘program’ T cell attraction to the epidermal chemokine CCL27. Nat Immunol. 2007; 8:285-93. [PubMed: 17259988]
Overberg L, Decallonne B, Valckx D, Verstuyf A, Depovere J, Laureys J, et al. Identification and immune regulation of 25-hydroxyvitamin D-1-alpha-hydroxylase in murine macrophages. Clin Exp Immunol. 2000; 120:139-46. [PubMed: 10759775]
Overberg L, Stoffels K, Waer M, Verstuyf A, Bouillon R, Mathieu C. Immune regulation of 25- hydroxyvitamin D-1 alpha-hydroxylase in human monocytic THP1 cells: mechanisms of interferon-gamma-mediated induction. J Clin Endocrinol Metab. 2006; 91:3566-74. [PubMed: 16787983]
Chawla A, Repa R, Evans M, Mangelsord DJ. Nuclear receptors and lipid physiology: opening the X-files. Science. 2001; 294:1866-70. [PubMed: 11729302]
Lin R, White JH. The pleiotropic actions of vitamin D. Bioessays. 2004; 26:21-8. [PubMed: 14696037]
Yasmin R, Williams RM, Xu M, Noy N. Nuclear import of the retinoid X receptor, the vitamin D receptor, and their mutual heterodimer. J Biol Chem. 2005; 280(48):40152-60. [PubMed: 16204233]
Takeuchi A, Reddy G, Kobayashi T, Okano T, Park J, Sharma S. Nuclear factor of activated T cells (NFAT) as a molecular target for 1,25-dihydroxyvitamin D3-mediated effects. J Immunol. 1998; 160:209-18. [PubMed: 9551973]
Lemire JM, Archer DC, Beck L, Spiegelberg HL. Immunosuppressive actions of 1,25- dihydroxyvitamin D3: preferential inhibition of Th1 functions. J Nutr. 1995; 125:1704S-8S. [PubMed: 7782931]
Van Etten E, Mathieu C. Immunoregulation by 1,25-dihydroxvitamin D3: basic concepts. J Steroid Biochem. 2005; 97:93-101.
Boonstra A, Barrat FJ, Crain C, Heath VL, Savelkoul HF, O’Garra A. 1-alpha-25- Dihydroxyvitamin D3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J Immunol. 2001; 167:4974-80. [PubMed: 11673504]
Liu PT, Krutzik SR, Modlin RL. Therapeutic implications of the TLR and VDR partnership.
Trends Mol Med. 2007; 13:117-24. [PubMed: 17276732]
Medzhitov R. Recognition of the microorganisms and activation of the immune response. Nature. 2007; 15:819-26. [PubMed: 17943118]
Saito T, Gale M. Principles of intracellular viral recognition. Cur Opin Immunol. 2007; 19:17-23.
Oberg F, Botlin J, Nilsson K. Functional antagonisms between vitamin-D3 and retinoic acid in the regulation of CD14 and CD23 expression during monocytic differentiation of U-937 cells. J Immunol. 1993; 150:3487-95. [PubMed: 7682243]
Daher K, Selsted M, Lehrer R. Direct inactivation of viruses by human granulocyte defensins. J Virol. 1986; 60:1068. [PubMed: 3023659]
Wang TT, Nestel FP, Bourdeau V, Nagai Y, Wang Q, Liao J, et al. Cutting edge: 1,25- dihydroxvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J Immunol. 2004; 173:2909-12. [PubMed: 15322146]
Liu PT, Schenk M, Walker VP, Dempsey PW, Kanchanapoomi M, Wheelwright M, et al. Convergence of IL-1beta and VDR activation pathways in human TLR2/1-induced antimicrobial responses. PLoS One. 2009; 4(6):e5810. [PubMed: 19503839]
Adams JS, Songyang R, Liu PT, Chun RF, Lagishetty V, Gombart AF, et al. Vitamin D-directed rheostatic regulation of monocyte antibacterial responses. J Immunol. 2009; 182:4289-95. [PubMed: 19299728]
Gombat AF, Borregard N, Koeffler HP. Human cathelicdin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor is strongly upregulated in myeloid cells by 1,25- dihydroxvitamin D3. FASEB. 2005; 19:1067-77.
Sorenson OE, Follin P, Johnsen A, Calafat J, Tjabringa G, Himestra PS. Human cathelicidin, hCAP-18 is processed to the antimicrobial peptide LL-37 by extracellular cleaveage with proteinase 3. Blood. 2001; 97:3951-9. [PubMed: 11389039]
Agerberth B, Charo J, Werr J, Olsson B, Idali F, Lindbom L, et al. The human antimicrobial and chemotactic peptides LL-37 and alpha-defensins are expressed by specific lymphocyte and monocyte populations. Blood. 2000; 96:3086-94. [PubMed: 11049988]
Zanetti M. The role of cathelicidins in the innate host defenses of mammals. Curr Issues Mol Biol. 2006; 7:179-96. [PubMed: 16053249]
Gordon YJ, Huang L, Romanowski E, Yates K, Proske R, McDermott A. Human Cathelicidin (LL-37), a multifunctional peptide is expressed by ocular surface epithelia and has potent antibacterial and antiviral activity. Cur Eye Res. 2005; 30(5):385-94.
Bals R, Wang X, Zasloff M, Wilson JM. The peptide antibiotic LL-37/hCAP-18 is expressed in the epithelia of the human lung where it has broad antimicrobial activity at the airway surface. Proc Natl Acad Sci USA. 1998; 95:9541-6. [PubMed: 9689116]
Chromek M, Slamova Z, Bergman P, Kovacs L, Pdracka L, Ehren I, et al. The antimicrobial peptide cathelicidin protects the urinary tract against the invasive bacterial infection. Nat Med. 2006; 12(6):636-41. [PubMed: 16751768]
Bals R, Wilson JM. Cathelicidins—a family of multifunctional antimicrobial peptides. Cell Mol Life Sci. 2003; 60:711-20. [PubMed: 12785718]
Howell MD, Jones JF, Kisich KO, Streib JE, Gallo RL, Leung DYM. Selective killing of vaccinia virus by LL-37: implications for eczema vaccinatum. J Immunol. 2004; 172:1763-7. [PubMed: 14734759]
Steinstraesser L, Tippler B, Mertens J, Lamme E, Homann HH, Lehnhardt M, et al. Inhibition of early steps in the lentiviral replication cycle by cathelicidin host defense peptides. Retrovirology. 2005; 2(1):1-12. [PubMed: 15644139]
Bergmana P, Walter-Jallow L, Broliden K, Agerberth B, Soderlund J. The antimicrobial peptide LL-37 inhibits HIV-1 replication. Curr HIV Res. 2007; 5(4):410-5. [PubMed: 17627504]
Hansdottir S, Monick MM, Hinde SL, Lovan N, Look DC, Hunninghake GW. Respiratory epithelial cells convert inactive vitamin D to its active form: potential effects on host defense. J Immunol. 2008; 181:7090-9. [PubMed: 18981129]
Hansdottir S, Monick MM, Lovan N, Powers L, Gerke A, Hunnighakw GW. Vitamin D decreases respiratory syncytial virus induction of NF-kB chemokines and cytokines in airway epithelium while maintaining the antiviral state. J Immunol. 2010; 184:965-74. [PubMed: 20008294]
Janssen R, Bont L, Siezen CL, Hodemakers HM, Ermers MJ, Doornbos G, et al. Genetic susceptibility to respiratory syncytial virus bronchiolitis is predominantly associated with innate immune genes. J Infect Dis. 2007; 196:826-34. [PubMed: 17703412]
Roth DE, Jones AD, Prosser C, Robison JL, Vohra S. Vitamin D receptor polymorphisms and the risk of acute lower respiratory tract infection in early childhood. J Infect Dis. 2008; 197:676-80. [PubMed: 18266602]
Rehman PK. Sub-clinical rickets and recurrent infection. J Trop Pedatria. 1994; 40:58.
Avenell A, Cook JA, Maclennan GS, Macpherson GC. Vitamin D supplementation to prevent infections: a sub-study of a randomized placebo-controlled trial in older people. Age Ageing.
2007; 36:574-7. [PubMed: 17702768]
Li-Ng M, Alolia JF, Pollack S, Cunha BA, Mikhail M, Yeh J, et al. A randomized control trial of vitamin D3 supplementation for the prevention of symptomatic upper respiratory tract infections. Epidemiol Infect. 2009; 137:1396-404. [PubMed: 19296870]
Vieth R, Chan PC, MacFralane GD. Efficacy and safety of vitamin D3 intake exceeding the lowest observed adverse effect level. Am J Clin Nutr. 2001; 73:288-94. [PubMed: 11157326]
Barger-Lux MJ, Heaney RP, Dowell S, Chen TC, Holick MF. Vitamin D and its major metabolities: serum levels after graded oral dosing in healthy men. Osteoporos Int. 1998; 8:22230. [PubMed: 9797906]
Laaksi I, Ruohola JP, Tuohimma P, Auvian A, Haatja R, Pilajamaki H, et al. An association of serum vitamin D concentrations < 40 nmol/L with acute respiratory tract infection in young Finnish men. Am J Clin Nutr. 2007; 86:714-7. [PubMed: 17823437]
Roth DE, Shah R, Baqui AH. Vitamin D status and acute lower respiratory infection in early childhood in Sylhet, Bangladesh. Acta Paediatri. 2010; 99:389-93.
Wayse V, Yousafzai A, Mogale K, Filteau S. Association of subclinical vitamin D deficiency with severe acute lower respiratory infection in Indian children under 5 y. Eur J Clin Nutr. 2004; 58:563-7. [PubMed: 15042122]
Karatekin G, Kaya A, Salihoglu O, Balci H, Nuhonglu A. Association of subclinical vitamin D deficiency in newborns with acute lower respiratory infection and their mothers. Eur J Clin Nutr. 2009; 63:473-7. [PubMed: 18030309]
McNally JD, Leis K, Matheson LA, Karuananyake C, Sankaran K, Rosenberg AM. Vitamin D deficiency in young children with severe acute lower respiratory infection. Pediatr Pulmonol.2009; 44:981-8. [PubMed: 19746437]
Roth DE, Jones AB, Prosser C, Robinson JL, Vohra S. Vitamin D status is not associated with the risk of hospitalization for acute bronchiolitis in early childhood. Eur J Clin Nutr. 2009; 63:297-9. [PubMed: 17971825]
Ginde AA, Mansbach JM, Camargo CA. Association between serum 25-hydroxyvitamin D level and upper respiratory tract infection in the Third National Health and Nutrition Examination Survey. Arch Intern Med. 2010; 169(4):384-90. [PubMed: 19237723]
Cannell JJ, Zasloff M, Garland CF, Scragg R, Giovannucci E. On the epidemiology of influenza. Virol J. 2008; 5:29. [PubMed: 18298852]
Urashima M, Segawa T, Okazaki M, Kurihara M, Wada Y, Ida H. Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Am J Clin Nutr. 2010; 91(5): 1255-60. [PubMed: 20219962]
Mayer J. Vitamin D and African Americans. J Nutr. 2006; 136(4):1126-9. [PubMed: 16549493]
Aloia JF, Li-Ng M. Re: epidemic influenza and vitamin D [with author reply]. Epidemiol Infect. 2007; 135:1095-8. [PubMed: 17352842]
Kuehn EW, Anders HJ, Bogner JR, OBermaier J, Goebel FD, Schlondorff D. Hypocalcaemia in HV infection and AIDS. J Intern Med. 1999; 245:69-73. [PubMed: 10095819]
Teichmann J, Stephan E, Lange U, Discher T, Friese, Lohmeyer J, Stracke H, Bretzel RG. Osteopenia in HIV-infected women prior to highly active antiretroviral therapy. J Infect. 2003; 46:221-7. [PubMed: 12799147]
Coodley GO, Codley MK, Nelson HD, Loveless MO. Micronutrient concentrations in the HIV wasting syndrome. AIDS. 1993; 7:1595-600. [PubMed: 7904452]
Haug C, Mueller F, Aukrust, Froland SS. Subnormal serum concentration of 1,25-vitamin D in human immunodeficiency virus infection: correlation with degree of immune deficiency and survival. J Infect Dis. 1994; 169:889-93. [PubMed: 7907645]
Dannhauser A, van Staden AM, van der Ryst E, Nel M, Erasmus E, Attwood EM, et al. Nutritional status of HIV-1 seropositive patients in the Free State Province of South Africa: anthropometric and dietary profile. Eur J Clin Nutr. 1999; 53:165-73. [PubMed: 10201796]
Villamor E. A potential role for vitamin D on HIV infection? Nutr Rev. 2006; 64(5):226-33. [PubMed: 16770943]
Cozzolino M, Vidal M, Arcidiacono MV, Tebas P, Yarasheski KE, Dusso AS. HIV-protease inhibitors impair vitamin D bioactivation to 1,25-dihydroxyvitamin D. AIDS. 2003; 17:513-20. [PubMed: 12598771]
Bang UC, Shakar SA, Hitz MF, Jespersen MS, Andersen O, Nielsen SD, et al. Deficiency of 25- hydroxyvitamin D in male HIV-positive patients: a descriptive cross-sectional study. Scand J Infect Dis. 2010; 42:306-10. [PubMed: 20085419]
Van Den Bout-Van Den Beukel CJP, Fievez L, Michels M, Sweep F, Hermus A, Bosch M, et al. Vitamin D deficiency among HIV type 1-infected individuals in the Netherlands: effects of antiretroviral therapy. AIDS Res Hum Retroviruses. 2008; 24(11):1375-82. [PubMed: 18928396]
Mueller NJ, Fux CA, Lederberg B, Elzi L, Schmid P, Dang T, et al. High prevalence of severe vitamin D deficiency in combined antiretroviral therapy-naive and successfully treated Swiss HIV patients. AIDS. 2010; 24:1-7. [PubMed: 19890206]
Rodriguez M, Daniels B, Gunawardene S, Robbins GK. High frequency of vitamin D deficiency in ambulatory HIV-positive patients. AIDS Res Hum Retroviruses. 2009; 25:9-14. [PubMed: 19108690]
Wasserman P, Rubin DS. Highly prevalent vitamin D deficiency and insufficiency in an urban cohort of HIV-infected men under care. AIDS Patient Care STDS. 2010; 24:223-7. [PubMed: 20377437]
Madeddu G, Spanu A, Solinas P, Calia GM, Lovingu C, Chess F, et al. Bone mass loss and vitamin D metabolism impairment in HIV patients receiving highly active antiretroviral therapy. O J Nucl Med Mol Imaging. 2004; 48:39-48.
Haug CJ, Aukrust P, Haug E, Morkrid L, Mueller F, Froland SS. Severe deficiency of 1,25- dihydroxyvitamin D3 in human immunodeficiency virus infection: association with immunological hyperactivity and only minor changes in calcium homeostasis. J Clin Endrocinol Metab. 1998; 83:3832-8.
de Luis DA, Bachiller P, Aller R, de Luis J, Izaola O, Terroba MC, et al. Relation among micronutrient intakes with CD4 count in HIV infected patients. Nutr Hosp. 2002; 17:285-9. [PubMed: 12514921]
O’Brien KO, Razavi M, Henderson RA, Caballero B, Ellis KJ. Bone mineral content in girls perinatally infected with HIV. Am J Clin Nutr. 2001; 73:821-6. [PubMed: 11273859]
Levin LI, Munger KL, Rubertone MV, Peck CA, Lennette ET, Spiegelman D, et al. Multiple sclerosis and Epstein-Barr virus. JAMA. 2003; 289:1533-6. [PubMed: 12672770]
Haahr S, Hollsberg P. Multiple sclerosis is linked to Epstein-Barr virus infection. Rev Med Virol. 2006; 16:297-310. [PubMed: 16927411]
Holmoy T, Vitamin. D status modulates the immune response to Epstein Barr virus: synergistic effect of risk factors in multiple sclerosis. Med Hypotheses. 2008; 70(1):66-9. [PubMed:17574770]
Bellamy R, Ruwende C, Corrah T, McAdam KPWJ, Thursz M, Whittle HC, et al. Tuberculosis and chronic hepatitis B virus infection in Africans and variation in the vitamin D receptor gene. J Infect Dis. 1999; 179:721-4. [PubMed: 9952386]
Ma J, Stampfer MJ, Gann PH, Hough HL, Giovannucci E, Kelsey KT, et al. Vitamin D receptor polymorphisms, circulating vitamin D metabolites, and risk of prostate cancer in United States physicians. Cancer Epidemiol Biomarkers Prev. 1998; 7:385-90. [PubMed: 9610787]
Tayeb MT, Clark C, Haites NE, Sharp L, Murray GI, McLeod HL. CYP3A4 and VDR gene polymorphisms and the risk of prostate cancer in men with benign prostate hyperplasia. Br J Cancer. 2003; 88:928-32. [PubMed: 12644831]
Ogunkolade BW, Boucher BJ, Prahl J, Bustin SA, Burrin JM, Noonan K, et al. Vitamin D receptor (VDR) mRNA and VDR protein levels in relation to vitamin D status, insulin secretory capacity, and VDR genotype in Bangladeshi Asians. Diabetes. 2002; 51:2294-300. [PubMed: 12086963]
Morrison NA, Qi JC, Tokita A, Kelley PJ, Crofts L, Nyguen TV, et al. Prediction of bone density from vitamin D receptor alleles. Nature. 1994; 367:284-7. [PubMed: 8161378]
Loke H, Bethell D, Phuong CX, Day N, White N, Farrar J, et al. Susceptibility to dengue hemorrhagic fever in Vietnam: evidence of an association with variation in the Vitamin D Receptor and Fc gamma receptor Ila genes. Am J Trop Med Hyg. 2002; 67(1):102-6. [PubMed: 12363051]
Sanchez-Valdezi E, Delgado-Aradillas M, Torres-Martinez JA, Torres-Benitez JM. Clinical response in patients with dengue fever to oral calcium plus vitamin D adminstration: study of 5 cases. Proc West Pharmacol Soc. 2009; 52:14-7. [PubMed: 22128411]
Bitetto D, Fabris C, Fornasiere E, Pipan C, Fumolo E, Cussigh A, et al. Vitamin D supplementation improves response to antiviral treatment for recurrent hepatitis C. Transpl Int. 201010.1111/j.1432-2277.2010.01141.x
Petta S, Camma C, Scazzone C, Tripodo C, Di Marco V, Bona A, et al. Low vitamin D serum level is related to severe fibrosis and low responsiveness to interferon-based therapy in genotype 1 chronic hepatitis C. Hepatology. 2010; 51(4):1158-67. [PubMed: 20162613]
Yano M, Ikeda M, Abe KI, Kawai Y, Kuroki M, Mori K, et al. Oxidative stress induces antihepatitis C virus status via the activation of extracellular signal-regualted kinase. Hepatology. 2009; 50:678-88. [PubMed: 19492433]
Leikina E, Delanoe-Ayari H, Melikov K, Cho MS, Chen A, Waring AJ, et al. Carbohydratebinding molecules inhibit viral fusion and entry by crosslinking membrane glycoproteins. Nat Immunol. 2005; 6(10):995-1001. [PubMed: 16155572]
Buck C, Day P, Thompson CD, Lubkowski J, Lu W, Lowy DR, Schiller JT. Human a-defensins block papillomavirus infection. Proc Nat Acad Sci USA. 2006; 103(5):1516-21. [PubMed: 16432216]
Kota S, Sabbah A, Chang TH, Harnack R, Xiang Y, Meng X, et al. Role of human a-defensin-2 during tumor necrosis factor-a/NF-kB-mediated innate antiviral response against human respiratory syncytial virus. J Biol Chem. 2008; 283:22417-29. [PubMed: 18567888]
626 citations as of Nov 2024
Google Scholar shows that this was cited by Google Scholar
Immunomodulatory role of vitamin D in infectious and non-infectious diseases - 2019 FREE PDF
Effect of high doses of vitamin D supplementation on dengue virus replication, Toll-like receptor expression, and cytokine profiles on dendritic cells - 2020 https://doi.org/10.1007/s11010-019-03658-w
The Potential Protective Role of Vitamin D Supplementation on HIV-1 Infection - Sept 2019 FREE PDF doi.org/10.3389/fimmu.2019.02291
Vitamin D in autoimmune, infectious and allergic diseases: A vital player? Aug 2011 DeepDyve
-
PLOS ONE June 2016, free PDF online
Transition probability to an epidemic state was 100% when < 2°C, < 4 g/m3 absolute humidity, and < 32 W/m2 solar radiation.
Vitamin D is a Secret Weapon Against Flu, COVID, and other enveloped viruses - Claude AI Aug 2025
Scientists have discovered why vitamin D works against some viruses but not others: it specifically targets the outer "envelope" that wraps around viruses like influenza, COVID-19, and RSV. This envelope acts like the virus's protective coat—and vitamin D produces tiny molecular scissors that cut right through it. The jaw-dropping evidence:84% flu reduction in kids taking vitamin D (but zero effect on non-enveloped viruses)
7% COVID reduction in healthcare workers on 4,000 IU daily
Your body literally manufactures antiviral peptides when vitamin D levels are optimal
The sweet spot: 1,000-4,000 IU daily (not massive monthly doses) targeting blood levels of 40-60 ng/mL—higher than most doctors recommend for bone health.
This isn't about boosting your immune system generally. It's about giving your body the raw materials to manufacture its own targeted antiviral weapons against some of the world's most dangerous respiratory viruses.
Exception: Measles is a rare enveloped virus which is not fought by Vitamin D
Wonder- Not enough vitamin D has been used?
- Might a vitamin D receptor activator help
See also VitaminDWiki
VitaminDWiki pages containing ENVELOPED in title 9 pages as of Oct 2021
Respiratory viral infection (RSV) and low vitamin D - many studies
Four antiviral strategies – includes vitamin D – Vasquez Dec 2014
Hepatitis B clinical event was 2X more likely if low vitamin D – Oct 2014
Vitamin D prevents Hepatitis-C and helps treat it (many studies)
Dengue fever - 8X higher risk for getting worse if low vitamin D – Sept 2017
Multiple Sclerosis etc. strongly associated with Epstein-Barr virus (and vitamin D)
Yet another virus associated with low vitamin D – mononucleosis – Aug 2016
Some enveloped virus are 1.2 X more likely if have a poor Vitamin D Receptor -Aug 2018
Long-haul fatigue, etc. common after viral infections (SARS1,2, MERS, Swine, 1918,...)
Rotavirus diarrhea (nearly every child gets it) is strongly associated with low vitamin D – Aug 2015
Vitamin D does not prevent all viral infections (enveloped virus not mentioned) – Jan 2019
Respiratory viral infection (RSV) and low vitamin D - many studies
LL-37 356 items as of June 2021
HIV category listing has items along with related searches
Immunity category listing has items along with related searches
See also web: Enveloped Virus protection
Curcumin from Turmeric Inhibits Zika, HIV, Herpes and Other Viruses GreenMedInfo April 2017
Reference includes the following
" Curcumin inhibits Zika and chikungunya virus infection by inhibiting cell binding" June 2017"
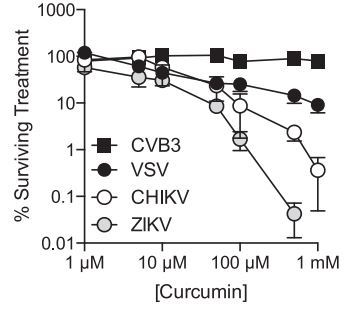
* Note: Curcumin activation of the Vitamin D Receptor increases amount of vitamin D getting into cells
* 📄 Download the PDF from VitaminDWiki
See also web
- Treating Ebola ebook Dr Sircus Sept 13, 2014, $20
- Using many things such as Sodium Bicarbonate, Vitamin D , Magnesium, Iodine, and very large amounts of vitamin C
- Virus chart from Wikipedia