Vitamin D Receptor role in Autoimmune Diseases and or cancers
Distilling the Genetics of the Vitamin D Receptor to Understand the Controversies over Beneficial Effects of Vitamin D in Autoimmune Diseases and/or Cancers
Human Genet Embryol 2013, 3:3 http://dx.doi.org/10.4172/2161-0436.1000e110
Yue Zhang zhanglee2006@gmail.com Department of Osteoarthritis Research Unit, University of Montreal Hospital Research Centre (CRCHUM), Notre-Dame Hospital, Montreal, Quebec H2L 4M1, Canada
Primer
Decoding the genetics of an increased risk of complex diseases such as Autoimmune Diseases (ADs) and/or cancers associated with the aging process remains challenging. The controversies exist for the beneficial effects of vitamin D supplementation on these diseases [1,2]. However, based on comparative advantages of different model systems and recent ChIP-seq/ChIP-chip studies, we distilled out one novel testable hypothesis as follows: the genetic regulatory network of vitamin D receptor (VDR, the homologue of Caenorhabditis elegans DAF-12) may play a central role as a common basis preventing some autoimmune diseases and associated cancers [3]. Further, as a capacitor, DAF-12/VDR may buffer polygenic genetic mutations and/ or variations. The highly-conserved targets (e.g. MMP3) of DAF-12/ VDR may have synergic functions with its other evolutionarily-"fresh" targets (e.g. IL-6). Being consistent with the "hygiene hypothesis" and the "danger signals" theory, some VDR/DAF-12 targets may be involved in these processes. In nature, the reversal of diseased status with vitamin D supplementation could be more difficult and/or complex than the breakdown of robust health. Put aggregates, they may partially clarify the controversies.
VDR and DAF-12 as a capacitor
All human beings need to buffer the influences of genetic variations and environmental challenges if we need to develop normally and be in good health. Furthermore, genes near these DAF-12/VDR binding sites network microRNA regulations, autophagy, longevity and cellular reprogramming, and so on. Probably similar to DAF-12, we all balance stability and the potential for change in part through the VDR receptor's involvement in developmental robustness [4]. Transient decreases in the VDR pathway resulting from of vitamin D deficiency, and/or a lack of UVB could uncover some morphological genetic mutations/ variants; possibly, mal-functional VDR genetically 'sensitizes' the pathways and destabilizes certain phenotypes (e.g. diseased status) and may reveal potent natural variations affecting these health phenotypes, i.e. disease.
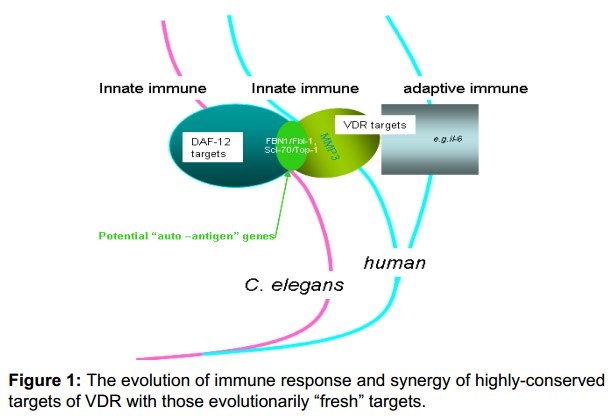
Figure 1 : The evolution of immune response and synergy of highly-conserved targets of VDR with those evolutionarily "fresh" targets.
Moreover, DAF-12/VDR may support both the activators and the inhibitors of the same function in a network which is full of feedback loops, and alteration of the DAF-12/VDR function could uncover variations that would adjust some processes either upwards or downwards [5]. The collective outputs are subjected to the whole network fine adjustments. Alternatively, genetic variation in the pathways of vitamin D metabolism, transport or signalling could modify the effects of vitamin D status on immunity to pathogens, as previously shown for tuberculosis [6]. Moreover, when DAF-12/VDR function is disturbed, developmental pathways or health maintenance programs are sensitized to a degree that is determined by their specific dependence on DAF-12/VDR by the functional significance and inherent stabilities of the relevant targets and by vitamin D availability. In this point, DAF-12/VDR may follow a similar model to the threshold trait previously designed for HSP90, which requires at least six genetic democratic determinants and assuming in population containing ten independent and additive determinants affecting the trait [5]. However, as in the case of HSP90, if a compromised DAF-12/VDR function was to lower the trait's threshold by just one or two determinants, the probability of the appearance of the trait increases 10- to 100-fold. Alongside adjustments of multiple signal transducers and thereby simultaneously disturbing several developmental pathways as DAF-12/VDR, mal-functional VDR may allow the threshold to be lowered, remodelling many different processes at once [7]. Further, once the appearance of a trait increases in this manner, for a moderate fitness advantage, selection could increase the frequency of genetic polymorphisms affecting the trait to a point at which it becomes independent of reduced DAF-12/VDR function. If this could take place, vitamin D supplements may not show the expected beneficial effects.
VDR and ADs
One Genome-Wide Association Study (GWAS) suggests that one allelic VDR variant may link to clinical autoimmune antibodies including anti-p150 (TRIM33/ TIF-1y)/p140(TRIM24/TIF-1a), whose natural self-/ auto-antigens may be related to protein products encoded by TIF-1y/a, their homologues flt-1 and nhl-2 as direct targets of DAF-12/VDR [8,9].

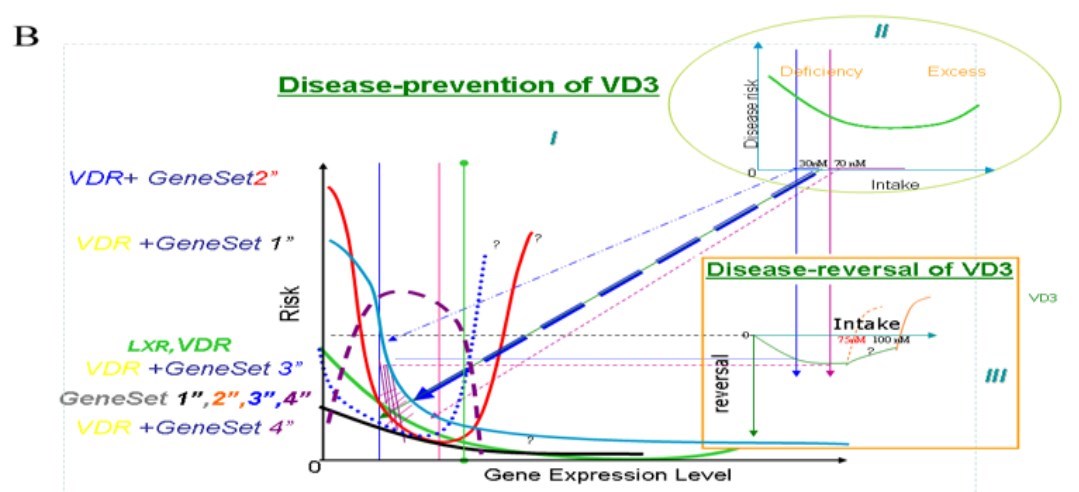
Figure 2 : Classifications of interactions between VDR and gene-sets along with disease risk and vitamin D intake.
I. The curve of disease risk and gene expression level. II. Disease risk and vitamin D intake. III. Risk reduction and vitamin D intakes.
Note: "?" means "unknown" and/or "not sure".
The grid represents the "buffering" capacity of VDR to risk of mal-function of other particular gene sets' genetic mutations.
Intake more than 140nM of VD3 may largely have "bad" effects. a: Type 1-Gene-Set 1: potentially a "standard" curve; it shows evident effects with a vitamin D intake less than 70 nM.
The risk curve for VDR+ Gene-Set 1 means the risk alongside deficiency in expression of both sets of genes.
Type 2-Gene-Set 2: A documented clinical cancer-related curve. An intake of <30 nM and > 70 nM intake has positive effects; but an intake of 30 nM-70 nM has negative effects. This group follows a U-shape [1,23,24]. However, somewhat consistently for this type, mutant animals have neither null alleles nor known loss/gain of function alleles, but only some animals with daf-12 RNAi feeding on DAF-12/VDR (rh274, gain of function) have a weak multi-vulval phenotype in our observations [25]. The multi-vulva may somewhat be considered an incomplete/imperfect cancer model in C. elegans.
Type3-Gene-Set 3: VDR has almost no effect at low vitamin D concentrations and intakes but becomes effective when vitamin D intake is more than 50 nM (or 70 nM in different study populations)
Type 4-Gene-Set 4: This has a very limited window of low vitamin D concentrations with positive effects; all other doses produce negative effects.
b: Type 5-Gene-Set 1" to Type 8-Gene-Set 4": VDR and vitamin D intake generally have positive effects.
Our ChIP-chip screening for DAF-12/VDR target genes overlap many validated homologues identified in human VDR studies and significantly enriched near genes that are pathologically associated with ADs and cancer. Another Genome-Wide Association Study (GWAS) identified genetic variants for joint damage progression in autoantibody-positive rheumatoid arthritis (RA) whose three key genes (Sperm-Associated Antigen 16 (SPAG16), and Matrix Metallopeptidase 1 and 3 (MMP1 and MMP3) are among human homologue candidates of DAF-12/vitamin D receptor (VDR) target genes, i.e. wdr-5.1 (with C. elegans cosmid gene name C14B1.4) and H36L18.1 (but there is single congruent C. elegans homologue for both human MMP1 and MMP3) correspondingly [4,7,10-13]. Uniquely, SPAG16 influences MMP3 activity and protects against joint destruction in auto-antibody-positive RA. Vitamin D3 is reported to control MMP3 in cultured human cells [14]. Consistently; a mal-function of VDR could affect the pathogenesis of RA and associated cancers [3,15]. Particularly, a better inhibition of human Th17- mediated synovial inflammation was obtained anti-TNF therapy alone than along with 1, 25(OH) 2D3-an active vitamin D metabolite; MMP3 made a contribution during this process. The highly-conserved targets of DAF-12/VDR (e.g. MMP3/MMP1) may possibly have synergistic functions with its other evolutionarily "fresh" targets (e.g. interleukin-6) (Figure 1) [7,8,16-18].
Novel Understanding of Controversies of Vitamin D Effects on Ads and Cancer
Vitamin D deficiency is highly prevalent in adults or children in northern America, Northern Europe and southern Asia, and has an association with susceptibility to diseases. Vitamin D may peak circulating concentrations of calcifediol, active vitamin D metabolite, which causes innate immune responses to microbes in vitro. The innate antimicrobial responses in vitro of calcifediol rely on the expression of receptors; consequently, vitamin D supplementation might have a better enhancement of immune function in one gene-set population than in others [19,20]. However, the Institute of Medicine initiated some "misconceptions" about vitamin D which have implications for clinicians and the role of vitamin D supplementation in patients with ADs [1,2,21]. Semira Manaseki-Holland et al. reported the results of a trial of bolus-dose vitamin D supplementation for disease prevention, but that report showed no beneficial effect [22]. Whether the vitamin D beneficial effects hypothesis is flawed is starting to be considered. The first possible interpretation is that the liver X receptor/LXR and VDR may share some or all of the set of targets of DAF-12/VDR so they are likely to be synergistic and have a comparable role to their counterpart DAF-12 in C. elegans. Moreover, their ligands may include Vitamin D3, Dafachronic Acid (DA) and possibly other micronutrients or environmental factors as well. Thus, vitamin D effects could be obscured by other micronutrients. Third, some responsive groups have a definitive gene-set but other non-responsive human groups have different gene-sets, which will somewhat weaken its beneficial effects. For simplicity, we hereby classify eight different types of potential interaction between VDR and different polygenic gene-sets (Figure 2). The different ADs may respond correspondingly with vitamin D supplementation. We speculate that the confusion would naturally arise along with some critical trials onto out-breeding populations like patients without precise classifications with gene-sets but also sparseness in experimental data at the whole organism system-level.
Type 1-polygenic Gene-Set 1
The standard disease curve frequently shows evident effects with an intake less than 70 nmol/L (nM). Protective effects might have been restricted to those with profound deficiency, as recently reported in a trial of vitamin D supplementation in human adults with chronic obstructive pulmonary disease.
Type 2-Gene-Set 2
A documented clinical cancer -related curve like. Intakes of <30 nM or >70 nM have significant positive effects; but intakes of 30 nM-70 nM have significant negative effects. This group follows a U-shape [1,23,24]. However, somewhat phenotypically consistent for this type, mutant animals have neither null alleles nor known loss/gain of function alleles, but only some animals with daf-12 RNAi feeding on DAF-12/VDR (rh274, gain of function) have a weak multi-vulval phenotype in our observations [25,26]. Note, the multi-vulva may possibly be considered one non-typical cancer model in C. elegans.
Type 3-Gene-Set 3
VDR has almost no effect at low vitamin D concentrations and intakes, but it becomes effective when the vitamin D intake is more than 50 nM (or 70 nM for some populations).
Type 4-Gene-Set 4
It has a very limited window of low concentrations of vitamin D with positive effects, and then all other doses produce negative effects. The peaks and troughs of circulating calcifediol concentrations could have bad effects on the immune response and it was observed that concentrations of calcifediol greater than 140 nM impaired the immunity to infection, possibly due to the suppression of vitamin D in adaptive responses to infection as well as boosting innate responses and thus recurrence of the disease [9,10,22].
Type 5-Gene-Set 1" to Type 8-Gene-Set 4"
VDR and Vitamin D intake generally have positive effects.
Most polygenic gene-sets have weaker (Types 1- 4) or no phenotype (Types 5-8); similarly, for VDR (and its friend LXR); but they may all make patients have an incompetent or predisposition status. For simplicity, Gene-Sets 1-4 and 1'-4' were put into the same curve. VDR (and LXR) was further simplified into either weaker or stronger phenotypes in comparison with theses gene-sets.
Of certain, the effects of vitamin D on its disease reversal would be weaker than its disease-prevention abilities in that the breakdown of robustness might be much easier than its rebuilding; thus, in many cases, multiple drug combinations may be expected to be used, given that a single vitamin D supplement is insufficient. In addition, if DAF-12/VDR buffering is compromised (for example, by a lack of UVB and vitamin D deficiency), cryptic disease variants are expressed and selection can lead to the continued expression of these traits, even when VDR function is restored with vitamin D supplements. Therefore a synthetic drug screen would need to be carried out. Besides, the results of individual studies cannot be exaggerated for its generalisability [10]. First of all, a variety of malnutrition/environmental factors will need to be taken into account in different study populations: some might therefore have been at high risk of deficiencies in other micronutrients such as calcium and vitamin A, many of which could make the beneficial effects of vitamin D supplementation or have some complementary effects (e.g. as seen in the counterpart for human dafachronic acid (DA) in C. elegans) to vitamin D supplementation [9,18]. As those with different nutritional backgrounds, caution should also be needed to extrapolate the results of one study to different gene-set populations. One subgroup with participants with a definitive gene-set might have benefited from vitamin D supplementation, but this effect was obscured by a larger group of less responsive participants [10]. In addition, after treatment with a certain dose of vitamin D, among multiple ADs, one or more hidden or mild diseases may be cured (but not the one initially targeted and then undetected) and many others may be improved (again, not targeted and undetected), but not many others. If the latter may be the most common situation in clinic trials up to now, superficially we cannot expect the evident beneficial effects to show up.
Dosing, Recommendations and Suggestions
Indeed, the negative outcome of any study is meaningful in that it possibly gives us the range of vitamin D at work and it informs the design of future studies [10]. A different dosing regimen of vitamin D in a different population might yield a positive result for a certain dose for disease prevention in one population with no beneficial effect.
Further, we may propose to assess effectiveness of vitamin D dosing in a population with high prevalence of deficiency and high incidence of disease such as Gene-Sets 1 and 2 (Figure 1a) as a logical point of departure [16]. Moreover, some nutritional or environmental factors may have synergic or complementary effects on vitamin D with clinical relevance. Another recommendation is to undertake trials of more frequent dosing regimens in other meticulously-designed age/geographical/ethnic groups with lower rates of malnutrition, characterising potential effect modifiers such as baseline vitamin D status and genetic factors [17]. In addition, being equipped with the complexity of organisms, different developmental stages of worms may be pooled as whole animals rather than using in vitro-focused mammalian cell lines, such screens are expected to cover VDR/DAF-12 target genes better. Further efforts using ChIP-seq on different stages with stresses, as well as proteomics assay, will shed further light on this direction. Importantly, C. elegans genome is much more compact than human genome so that the associate loci do tell us immediately downstream target candidates 4. Thorough understanding the underlying mechanisms may reduce the "contribution" of "shot-gun" content in clinical trials.
Conclusions
In summary, this essay is a primer for the current status of the genetic regulatory network of VDR (and DAF-12) and vitamin D supplementation in protection from ADs and/or associated cancers. Probably, therapeutic supplementation with a definitive range of systematically optimized dosages of vitamin D would need to explore how to maximize its beneficial effects within some combined treatments of such diseases. If this can be proven true, Mendel genetics could be evolved for complex diseases as well.
References
1. Rosen CJ, Taylor CL (2013) Common misconceptions about vitamin D--implications for clinicians. Nat Rev Endocrinol 9: 434-438.
2. White JH (2013) Vitamin D and human health: more than just bone. Nat Rev Endocrinol 9: 623.
3. Zhang Y (2013) Genetic basis of DAF-12/vitamin D receptor (VDR) in autoimmune immunity, autoimmune diseases and associated cancers. Clon Transgen 2: e105.
4. Greer EL, Maures TJ, Hauswirth AG, Green EM, Leeman, et al. (2010) Members of the H3K4 trimethylation complex regulate lifespan in a germline-dependent manner in C. elegans. Nature 466: 383-387.
5. Rutherford SL, Lindquist S (2010) Hsp90 as a capacitor for morphological evolution. Nature 396: 336-342.
6. Martineau AR, Leandro AC, Anderson ST, Newton SM, Wilkinson KA, et al. (2010) Association between Gc genotype and susceptibility to TB is dependent on vitamin D status. Eur Respir 35: 1106-1112.
7. Hochbaum D, Zhang Y, Stuckenholz C, Labhart P, Alexiadis V, et al. (2011) DAF-12 regulates a connected network of genes to ensure robust developmental decisions. PLoS Genet 7: e1002179.
8. Kapoor S, Cooper S, Robert G, Chinoy, Hector, et al. (2009) The Relationship Between the Vitamin D Receptor Gene and Anti-155/140 Antibodies in UK Caucasians with Idiopathic Inflammatory Myositis [abstract]. Arthritis Rheumatism 60: 805.
9. Nielsen NO, Skifte T, Andersson M, Wohlfahrt J, Soborg B, et al. (2010) Both high and low serum vitamin D concentrations are associated with tuberculosis: a case-control study in Greenland. Br J Nutr 4: 1487-1491.
10. Martineau AR (2010) Bolus-dose vitamin D and prevention of childhood pneumonia. Lancet 379: 1373-1375.
11. Knevel R, Klein K, Somers K, Ospelt C, Houwing-Duistermaat JJ, et al. (2013) Identification of a genetic variant for joint damage progression in autoantibody-positive rheumatoid arthritis. Ann Rheu Dis.
12. Motola DL, Cummins CL, Rottiers V, Sharma KK, Li T, et al. (2006) Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 124: 1209-1223.
13. Yang CY, Leung PS, Adamopoulos IE, Gershwin ME (2013) The Implication of Vitamin D and Autoimmunity: a Comprehensive Review. Clin Reviews Allergy Immunol 45: 217-226.
14. Schmitz JP, Schwartz Z, Sylvia VL, Dean DD, Calderon F, et al. (1996) Vitamin D3 regulation of stromelysin-1 (MMP-3) in chondrocyte cultures is mediated by protein kinase C. J Cell Physiol 168: 570-579.
15. Zhang Y (2013) Gaps! Transgenesis, model organisms, and human diseases. Clon Tansgen 2: e103.
16. Plenge RM, Greenberg JD, Mangravite LM, Derry JM, Stahl EA, et al. (2013) Crowdsourcing genetic prediction of clinical utility in the Rheumatoid Arthritis Responder Challenge. Nat Genet 45: 468-469.
17. Van Hamburg JP, Asmawidjaja PS, Davelaar N, Mus AM, Cornelissen F, et al. (2012) TNF blockade requires 1,25(OH)2D3 to control human Th17-mediated synovial inflammation. Ann Rheu Dis 71: 606-612.
18. Noyola-Martinez N, Diaz L, Avila E, Halhali A, Larrea F, et al. (2013) Calcitriol downregulates TNF-alpha and IL-6 expression in cultured placental cells from preeclamptic women. Cytokine 61: 245-250.
19. Black RE, Cousens S, Johnson HL, Lawn JE, Rudan I, et al. (2010) Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet 375: 1969-1987.
20. Liu PT, Stenger S, Li H, Wenzel L, Tan BH, et al. (2006) Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 311: 1770-1773.
21. Abrahamsen B, Harvey NC (2013) The role of vitamin D supplementation in patients with rheumatic diseases. Nat Rev Rheumatol 9: 411-422.
22. Manaseki-Holland S, Maroof Z, Bruce J, Mughal MZ, Masher MI, et al. (2012) Effect on the incidence of pneumonia of vitamin D supplementation by quarterly bolus dose to infants in Kabul: a randomised controlled superiority trial. Lancet 379: 1419-1427.
23. Grant WB (2009) Critique of the U-shaped serum 25-hydroxyvitamin D level-disease response relation. Dermatoendocrinol 1: 289-293.
24. Ramos PS, Criswell LA, Moser KL, Comeau ME, Williams AH, et al. (2011) a comprehensive analysis of shared loci between systemic lupus erythematosus (SLE) and sixteen autoimmune diseases reveals limited genetic overlap. PLoS Genet : e1002406.
25. Zhang Y (2011) Biology of the Mi-2/NuRD Complex in SLAC (Stemness, Longevity/Ageing, and Cancer). Gene Regul Syst Bio 5: 1-26.
26. Lehouck A, Mathieu C, Carremans C, Baeke F, Verhaegen J, et al. (2012) High doses of vitamin D to reduce exacerbations in chronic obstructive pulmonary disease: a randomized trial. Ann Inter Med 156: 105-114.
I have very little understanding of what this paper is attempting to say. Henry Lahore, admin of VitaminDWiki
See also VitaminDWiki
Genetics items
Vitamin D receptor polymorphisms are risk factors for various cancers – meta-analysis Jan 2014
The items in both Autoimmune and VDR are:
{category}
Vitamin D Receptor is associated in over 40 autoimmune studies
Vitamin D Receptor category items
| ** | ||
|---|---|---|
| Increased Risk | Cancer (Oct 2017) | |
| 4.6 | Breast Cancer | |
| 3.1 | Colon Cancer survival | |
| 2.7 | Gastric Cancer | |
| 2.4 | Lung Cancer | |
| 2 | Melanoma | |
| 1.6 | Prostate Cancer while black | ** |