Malignant melanoma may be reduced by skin-activated vitamin D
Current knowledge on the active form of Vitamin D synthesized in the skin and its effects on malignant melanoma
Neoplasma, doi:10.4149/neo2017101, Received April 18, 2016 / Accepted August 27, 2016
b. bolerazska beata@moly.sk, m. rabajdova, i. spakova, m. marekova
Department of Medical and Clinical Biochemistry, Faculty of Medicine, P. J. Safarik University in Kosice, Kosice, Slovakia
 ---
VitaminDWiki pages with MELANOMA in title (41 as of Jan 2022)
This list is automatically updated
{LIST()}
---
VitaminDWiki pages with MELANOMA in title (41 as of Jan 2022)
This list is automatically updated
{LIST()}
📄 Download the PDF from VitaminDWiki
The link between sunlight and skin cancer is a frequently discussed topic. However, ultraviolet radiation also induces the production of vitamin D in the body. Keratinocytes and their ability to synthesize the active form of vitamin D, which is consumed at the place of its origin in the skin, have a unique place in this discussion. We observe a remarkable sunshine- related paradox when we monitor the relationship between the dose of solar radiation and one type of skin cancer - malignant melanoma. Recent knowledge of the non-calcemic effects of vitamin D, which include growth regulation, DNA repair, differentiation, apoptosis, membrane transport, metabolism, cell adhesion and oxidative stress, could help to further clarify this relationship. In this context, adjuvant vitamin D therapy is currently being considered in patients with malignant melanoma, and this is expected to reduce tumor invasiveness and micrometastases and thus improve patient prognosis and reduce the risk of relapse.
Over the last decade the so-called “sunshine vitamin” has again become a hot topic of discussion. The reasons for this are the recently gained knowledge on its noncalcemic effects in humans and the finding that serum levels of vitamin D, based on the results of epidemiological studies and current physiologic serum levels, are globally considered to be insufficient and are being referred to as a pandemic of vitamin D deficiency [1]. In the context of this pandemic and the skin synthesis of the active form of vitamin D, the following questions are raised: What effect do the reduced levels of the active form of vitamin D or its precursors have on metabolism in the skin? Could the skin be selfsufficient in the production of vitamin D? Can exogenous vitamin D supplements affect the risk of developing skin diseases, in particular skin cancer? Answering these questions clearly and responsibly is not currently possible. In this article we try to summarize the latest knowledge and the relationship between the unique synthesis of the active form of vitamin D in the skin directly and its impact on the disease malignant melanoma (MM). The basic terminology of vitamin D is shown in Table 1.
Localization of endogenous Vitamin D synthesis
The generally known process of human endogenous vitamin D synthesis begins in the skin after exposure to Uv radiation, and its conversion to the active form occurs by hydroxylation in the liver and kidneys. A prerequisite for the course of such hydroxylation reactions is the presence of the enzymes 25-hydroxylase (CYP27A1) and 1a-hydroxylase (CYP27B1); the first enzyme is mainly active in hepatocytes and the second mainly in the proximal tubule of the kidneys. The 25(OH)D3 generated by the photochemical reaction represents about 90% of the circulating vitamin D level. The other 10% comes from food sources and is referred to as ergocalciferol (25(OH)D2).
Table 1. The basic terminology of Vitamin D
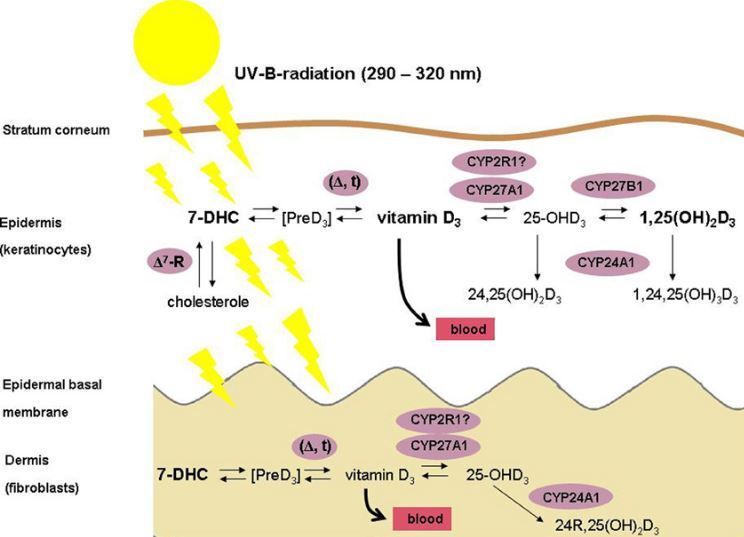
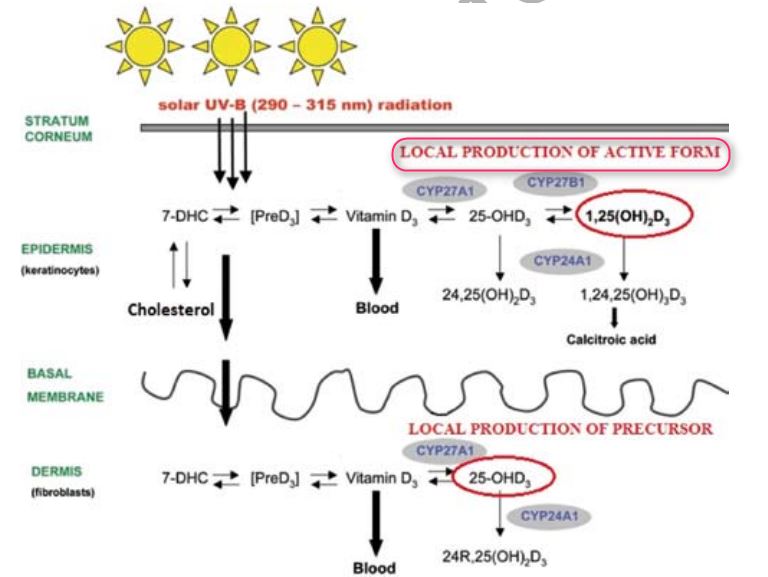
Less well known is the fact that this synthesis also runs simultaneously and in its entirety in keratinocytes, which are the only human cells with the enzymes needed for the full course of the synthesis from pro-vitamin vitamin D (7-dehydrocholesterol) to its active form (calcitriol, 1,25(OH)2D3) (Figure 1) . similarly, the significance and function of the active vitamin D formed in this way is less clear [2]. Upon learning these facts, efforts have been made to assess how much active vitamin D (1,25(oH)2D3) the skin is able to synthesize.
Extra-renal synthesis of 1,25(oH)2D has been repeatedly demonstrated in anephric humans [3]. At present it is not yet clear how much of the 1,25(oH)2D3 is produced by the epidermis and actually enters into circulation. experiments for these purposes measured the production of the active form of vitamin D in the skin of pigs without kidneys and evaluated the extent to which this participates in the total production of 1,25(oH)2D3 in systemic circulation in the basal state and after supplementation with precursors of the active form of Vitamin D. It was shown that only a minimal amount of the Vitamin D in systemic circulation was formed by the skin in the active form under normal conditions, but in anephric animals this proportion is increased [4]. UVB - triggered synthesis of calcitriol in human skin was demonstrated for the first time in vivo only in 2003 [5]. This means that 25(OH)D of photochemical origin, although predominant in systemic circulation, is not completely released from the skin, but a certain fraction is subjected to progressive hydroxylation directly in the keratinocytes. Fibroblasts in the skin can be a natural donor of precursors of the active form of Vitamin D, because they produce one of the necessary enzymes, 25-hydroxylase (CYP27A1), but not the other, 1a-hydroxylase (CYP27B1) [6]. The cutaneous synthesis of calcitriol (1,25(OH)2D3) is attributed to the effect of intracrine and/or autocrine on the keratinocytes themselves and paracrine on the adjacent cells, which may be melanocytes or malignant melanoma cells [7].
In the blood, the precursor of the active form (25(OH) D) is almost completely bound to the Vitamin D binding protein (VDBP); only about 0.3% is free [8]. The result of this is that keratinocytes cannot utilize precursors of the active form of Vitamin D in systemic circulation and are almost entirely dependent on their own synthesis. The absence of blood supply to the epidermis also contributes to this lower utilization. Similarly, only 0.4% of the active form of Vitamin D in systemic circulation is in its free form [9]. According to the "free hormone hypothesis", it is generally accepted that it is only the free 1,25(OH)2D3, not the total amount, that controls the genomic process in keratinocytes [10]. However, its concentration in systemic circulation is far too low to induce the effects of the hormone mediated by the Vitamin D receptor (VDR) in the skin, although keratinocytes possess the VDR [11,12]. This is probably one of the reasons why concentrations of Vitamin D that are too high can affect metabolic processes in the skin (for example the inhibition of proliferation; stimulation of differentiation, including formation of the permeability barrier; promotion of innate immunity; and promotion of the hair follicle cycle [13], including carcinogenesis [14].
General biological effects of Vitamin D in the context of malignant melanoma
The general biological effects of Vitamin D in the context of cancer, malignant melanoma in particular, may occur at the tumor location and by passing into systemic circulation may also affect a tumor’s ability to form micrometastases. The remote and local effects of Vitamin D are mainly mediated by the VDR. This is a predominantly nuclear protein which binds the active form of Vitamin D with high affinity and specificity and in turn regulates the transcription of many genes: 1,25(OH)2D may regulate at least 3000 genes in the human genome [15]. In addition, the sensitivity of melanoma to added 1,25-dihydroxyVitamin D3 seems to correlate with the stimulation of gene expression. For example, MeWo and SK Mel-28 melanomas were found to be sensitive to 1,25-dihydroxyVitamin D3, which also resulted in altered expression of Vitamin D-related genes. In other melanomas, such as SK Mel 5 and SK Mel 25, treatment with 1,25-dihydroxyVitamin D3 failed to induce expression of the genes and inhibition of cell growth [16].
Modulation of the target genes is carried out after the binding of calcitriol to the VDR. Interaction of the RXR (Retinoid X receptor) to the ligand-binding domain on the VDR, the structurally conserved DNA part, is necessary for the formation of this bond. A complex consisting of the VDR, RXR and calcitriol is ready for binding to Vitamin D responsive elements (VDREs), regions on the DNA at various distances from the transcription start site of the gene being regulated [17]. The highest expression of this receptor has been reported in metabolically active tissues, such as the skin, intestine, kidney and thyroid. It is also expressed in tumor tissues [18]. Based on an immune-histochemical analysis of skin tissues, it was proven that expression of the VDR decreases in the following order: normal skin > melanocytic nevi > non-metastatic melanoma = metastatic melanoma. Likewise VDR expression decreases with increasing progression of the tumor stage [19]. This receptor is also exprimated by keratinocytes to make it possible to respond to their own product - the active form of Vitamin D. Moreover, we also know that the non-genomic actions of 1,25(OH)2D3 lead to the activation of many signaling molecules, such as phospholipase C, phospholipase A2 (PLA2), phosphatidylinositol-3 kinase (PI3K) and p21ras, and the rapid generation of second messengers (Ca2+, cyclic AMP, fatty acids and 3-phosphoinositides, such as phosphatidylinositol 3,4,5 trisphosphate), accompanied by the activation of protein kinases, such as protein kinase A, src, mitogen-activated protein (MAP) kinases, protein kinase C (PKC) and Ca2+-calmodulin kinase II [20, 21, 22, 23, 24].
The most important mediated biological anti-cancer effects of 1,25(OH)2D3 include the induction of cell-cycle arrest, stimulation of apoptosis and inhibition of metastasis and angiogenesis [25]. The major antiproliferative effect of 1,25(OH)2D3 is based on blocking the G1 phase of the cell cycle [26]. The up-regulation of p21 and p27 principally mediate G1 cell-cycle arrest, but in addition 1,25(OH)2D3 has also been shown to mediate G2/M cell-cycle arrest in a number of cancer cell lines [27, 28]. In general, Vitamin D can the affect cell cycle in several ways, for example, through expression of the cyclins D1E and A, the kinases CDK 2, 4 and 6 [29], and the proteins Myc, Fos, Jun; In addition, it can up regulate insulin-like growth factor binding protein-3 (IGFBP3) [30], decreate and degradate prostaglandins [31] and influence the phosphorylation of retinoblastoma protein [29]. VDR loss or loss of the ability to form 1,25(OH)2D3 (CYP27B1 mutations/ deletions) indirectly confirm the effects of Vitamin D, which in skin results in disruption of the epidermal differentiation process (in the epidermis), resulting in hyperproliferation of the basal layer. These findings have been proven in a relatively short period [32].
In the development of malignant melanoma the inflammatory reaction, which most often results in response to ongoing tumorigenesis or after exposure to UV radiation, plays an important role. Cancer-related inflammation is in general characterized by the presence of inflammatory cells at tumor sites and over expression of inflammatory mediators, such as cytokines, chemokines, prostaglandins (PCs) and reactive oxygen and nitrogen species, in tumor tissue [33, 34]. Calcitriol affects the PC pathway in general by three separate mechanisms: decreasing COX-2 expression, increasing 15-PDCH expression (an enzyme regarded as a physiological antagonist of COX-2) and reducing PC receptor levels [35, 36]. Promotion of tumor angiogenesis, metastasis and invasion may be due to activate angiogenic switches under the control of vascular endothelial growth factor (VCEF) by these above-mentioned mediators [37, 38]. Hypoxia significantly increases its production and it has been noted that Vitamin D is able to reduce VECF expression during exposure to hypoxic conditions [39, 40]. Vitamin D can inhibit the expression of VECF by cancer cells and decrease responses to VECF by endothelial cell [39, 41]. The anti-inflammatory effects of Vitamin D can be mediated through up regulation of the expression of mitogen- activated protein kinase phosphatase-5 (MKP5), which in turn reduces the level of expression of pro-inflammatory cytokines and also their biological activity [31]. Incorrect regulation of NF-kB, a known protein complex that among other things controls DNA transcription, cytokine production and cell survival, has been repeatedly linked to cancer development, notably in the process leading from inflammation to carcinogenesis [42]. In contrast to normal cells, many cancer cells have elevated levels of active NFkB. Calcitriol is able to block NFkB activation [43], and inhibition of NF-kB activation appears to be a very promising option for anti-cancer therapies, including for melanoma [44].
Vitamin D can induce the apoptosis of cancer cells at the gene level through the inhibition of the anti-apoptotic gene Bcl-2 [45] and the induction of pro-apoptotic genes such as DAP (death-associated protein-3), CFKAR (cyspase 8 apoptosis-related cystein peptidase) and FADD (Fas-associated death domain) [46] and can actually stimulate the pro-autophagic gene beclin-1 [47]. In addition, calcitriol enhances activation of the pro-apoptotic proteins Bax and (r-calpain [48, 49, 50, 51]. The ability to activate apoptosis is definitely one of the most important functions of tumor protein p53, and disruption of this process can promote tumor progression and chemoresist- ance. Protein p53 serves as a regulator of the apoptotic process that can modulate key control points in both extrinsic and intrinsic pathways. It has been shown in several cancer cell lines that the mechanism of Vitamin D-induced apoptosis varies with the cell type and can be mediated by both the p53-dependent as well as independent pathways [52, 53]. The functional convergence between p53 family and VDR signaling, which occurs in the dermis, is probably an evolutionary adaptation to counterbalance the conflicting physiological requirements of Vitamin D synthesis and genome protection to protect against genotoxic insults derived from either the environment or local inflammation [54].
Position of Vitamin D in the tumor microenvironment of malignant melanoma
The melanoma microenvironment includes principally the endothelium, inflammatory cells and keratinocytes. Under normal tissue homeostasis, melanocytes in the skin dwell on the basement membrane in close contact with keratinocytes, which direct their behavior and growth through an intricate system of growth factors and cell-adhesion molecules [55]. Keratinocytes are clearly involved in crosstalk with malignant melanocytes. These interactions between keratinocytes and melanocytes in relation to the local production of the active form of Vitamin D have not yet been examined in detail. Similar to keratinocytes, the autonomous local production of the active form of Vitamin D (1,25(OH)2D3) and also expression of the VDR [56] have also been found to occur in melanocytes. Melanoma cells are also capable of synthesizing 1,25(OH)2D3 from 25(OH)D3 and expressing the VDR, and on exposure to 1,25(OH)2D3 they respond by slowing their proliferation [57]. Both types of cells lack the complete enzymatic equipment for total synthesis of Vitamin D, and therefore they are dependent on the supply of intermediates or the active product directly from their surrounding environment. Data obtained from in vitro experiments proved that the active form of Vitamin D (1,25(OH)2D3) was able to stimulate the maturation of melanocytes, presumably through stimulation of tyrosinase activity (a key enzyme in melanin biosynthesis) [58, 59]. It also protected the cells from apoptosis and increased expression of the VDR [60, 61].
The multistep process that leads to neoplastic transformation includes genome instability, avoiding immune attack, evading growth suppressors, enabling replicative immortality, resisting cell death, sustaining proliferative signaling (including angiogenesis), activating invasion and metastasis and deregulating cellular energetics and tumor promoting inflammation [62]. The response to inflammation is pro-oxidant, with production of reactive oxygen species (ROS) and reactive nitrogen intermediates (RNI). The result is redox dysregulation, which promotes alteration in the signaling and leads to secretion of chemokines, cytokines and prostaglandins related to the onset of neoplasia [63]. Most studies are performed in vitro by growing isolated melanoma cells in monocultures under conditions that cannot accurately imitate the appropriate tumor microenvironment [64]. In some, but not all melanoma cell cultures, anti-proliferative and pro-differentiative effects of Vitamin D and its precursors were demonstrated [57, 65, 66]. The active form of Vitamin D (1,25(OH)2D3) inhibited the invasive behavior of tumor and angiogenesis in melanoma cell lines [67] and also suppressed the growth of human melanoma transferred as xenografts to immunosuppressed mice that express the vDR. However, this did not occur in MM cell lines, which were modified to not express the vDR [68]. subsequently, it was shown that certain melanoma cell lines were resistant to the effects of Vitamin D. These cell lines exhibited a decreased expression of mRNA for the gene encoding the vDR or increased activity of the enzyme 24-hydroxylase (CYp24A1), which is an enzyme regulating excess of Vitamin D [56, 66, 69]. Melanoma cell activity is, in addition to other factors, dependent on the activity of tumor-infiltrating inflammatory cells and fibroblasts. The melanoma cells are able to influence the differentiation pattern of keratinocytes by production of FGF-2, VEGE A, H-8, and CXd-1. The reciprocal activity of keratinocytes to melanoma cells needs further research [70]. The potential role of melanoma cell-activated keratinocytes on tumor biology, including metastasation, should be verified [70].
A long time ago it was assumed that mortality due to cancer could be reduced by mild unprotected exposure to UV radiation, or by oral substitution of Vitamin D3 [71]. Exposure to ultraviolet light results in augmented Th-1, H-6, IL-8 and TNF-a production by human keratinocytes [72, 73]. Topical application of 1,25(OH)3D3 to the UV-irradiated skin of a human subject reduced the "sunburn cells" numbers [74]. The term "sunburn cells" is used for apoptotic keratinocytes with a pyknotic nucleus and eosinophilic cytoplasm [75]. One of the demonstrated abilities of Vitamin D is optimization of DNA repair, which protects against UV-induced mutations, the most common cause of skin cancer in humans. A number of epidemiological studies have shown a link between Vitamin D status and different types of cancers [76, 77], and some of them have confirmed this hypothesis, showing the protective effect of Vitamin D against progression and overall mortality in a large group of different cancer types [78]. Interestingly, since UV exposure is the main risk factor for melanoma, Vitamin D synthesis associated with UV exposure may also serve as a protective factor [79, 80].
Behind all this hides the Sun
Life on Earth originated in harmony with the sun, and therefore Vitamin D should be considered as one of the oldest hormones evolutionarily . It is photosynthesized in all forms of life, from phytoplankton (750 million years ago) to mammals. While Vitamin D’s role in calcium and bone metabolism makes it clear why terrestrial animals need it, it is less clear why marine and fresh water invertebrates and plants generate Vitamin D [81]. UV light from sun exposure has several well-known effects in the skin: UVA induces DNA damage through increasing the level of reactive oxygen species (ROS), but importantly UVB light also catalyzes the conversion of 7-dehydrocholesterol to 25(OH)-D and even induces the expression of VDR . VDR probably represent an adaptation of the skin to UV exposure, coupling the paramount importance of initiating 1,25(OH)2D3 synthesis with protection of cell and tissue integrity [82]. Thus, VDR actions are able to maximize UV-initiated synthesis of 1,25(OH)2D3, whilst controlling the extent of local inflammation that can result from sun exposure. Inflamed tissues contain more ROS, which in turn can damage DNA and prevent the proper function of DNA-repair machinery. Also, the induction of cytokines and growth factors associated with inflammation act to increase the proliferative potential of the cells. The above-mentioned NF-kB - a key mediator of inflammation - and the VDR attenuate this process by negatively regulating NF-kB signaling [83]. The normally protective role of inflammation that occurs under other conditions is lost through VDR-mediated suppression, but it is compensated for by the induction of a cohort of antimicrobial and antifungal genes. The induction of antimicrobials not only prevents infection in damaged tissue but can be cytotoxic for cells with increased levels of anion phospholipids within their membranes, a common feature of transformed cells [84]. The effects of 1,25(OH)2D3 have been expanded to include its impact on nucleotide excision repair (NER), the main system of DNA repair, which is induced after exposure to ultraviolet light. Recognition of the damage with the help of specific ("Damage Sensing") proteins leads to the removal of the short, single-stranded DNA segment that contains the lesion. Then, as part of the NER mechanism, the synthesis of a complementary strand of DNA and ligation follow [85]. It is likely that 1,25(OH)2D3 can stimulate NER, leading to more efficient removal of carcinogenic UV-induced photoproducts and other lesions involved in skin cancer transformation [86]. In in vivo studies in hairless mice (Skh: HR1) with skin cancer induced by UV radiation, topical treatment with 1,25(OH)2D3 post-exposure appeared to reduce the amount of DNA damage as measured by the number of cyclobutane pyrimidine dimers (CPDs) formed [87]. A proposal to implement individualized administration of Vitamin D based on the analysis of the NER system for the purpose of prevention/treatment of skin cancers in today's world of personalized medicine has also emerged in this area. It has been noted by Pawlovska et al. (2016) and is based on isolation of keratinocytes from an individual and subsequent analyzing of the NER system (functional assay), on the basis of which the dosage of Vitamin D3 supplementation would be determined [86]. UV radiation also causes degradation of generated Vitamin D, and therefore it has a regulatory effect on its creation while almost completely eliminating the possibility of a Vitamin D overdose caused by sun exposure [88]. The mere formation of Vitamin D is influenced by numerous internal (e.g. skin thickness with its associated quantity of precursor, ethnicity, congenital enzyme activity) and external factors (e.g. way of dressing, geographic location, number of sunny days a year), but UV rays have the most significant impact on its creation, namely by photodegradation [89].
The most widespread current hypothesis on the origin of malignant melanoma (MM) is its development as a result of sunburning in people who spend the vast majority of time indoors, and who during a holiday are subjected to excessive sun exposure [90, 91, 92, 93]. This results in local damage to DNA and immunosuppression, which leads to an increased risk of malignant melanoma after some time on the intermittently covered parts of the body [94, 95]. A rising incidence of malignant melanoma is being continuously recorded globally, but this trend seems to be closely related to screening campaigns in the interests of public health, and their implementation has contributed to an artificial increase in the incidence of the disease in many countries. This means that changes in behavior towards sunlight are not one of the main causes of the increased incidence. Another argument that weakens the harmful effect of Uv light in connection with malignant melanoma is that melanoma is a tumor that behaves similarly within similar ethnic groups worldwide, despite the varying intensity of Uv radiation. The anticipated reduction in the average age of onset of the disease has not occurred and no changes have been recorded even in the most common localizations of malignant melanoma [96].
The incidence of MM on skin intermittently exposed to sunlight is significantly less common in people who work outdoors than in people working indoors [97]. This finding may mean a lower probability of sunburn in humans who are often exposed to the sun, but an alternative hypothesis is that these people are less likely to be vitamin D deficient. Another aspect being considered is the different pathogenesis of melanomas which occur in people with a higher risk of actinic skin damage (skin phototype). In 2010 an extensive study was conducted in the Uk whose results showed a more frequent occurrence of malignant melanoma in phenotypes with a tendency to sunburn and confirmed skin burning before the age of 20 [93]. In contrast, another current hypothesis regarding malignant melanoma is risk reduction due to photoadaptation after periodic exposure to solar radiation, which simultaneously increases the synthesis of vitamin D. some data even indicate a possible photoprotective effect of vitamin D itself [54, 74, 98]. vitamin D could be a mediator through which sunburn results in systemic immunosuppression [98]. vitamin D was found to cause a reduction in UvA-induced skin damage and also a reduction in Uv-induced immunosuppression in studies performed on mice models in vitro and also on human skin [99]. These effects of vitamin D were also demonstrated when vitamin D analogues were applied topically to irradiated skin [74].
In 2005, a study was published which monitored the presence of elastosis (dermal post-solar skin damage) in samples of malignant melanoma. The results indicated that the presence of dermal damage to the skin in the excision of malignant melanoma meant a better prognosis for the patient [100]. one explanation for this finding is that chronic sun damage can cause a less aggressive form of malignant melanoma. Another possible explanation is that the presence of higher levels of vitamin D in individuals exposed to the sun more often can protect against the formation of relapse, even in terms of the link between Uv radiation and the etiology of this disease [101].
Thus, if we assume that the anti-proliferative effect of vitamin D is important for modifying the development of the disease in patients with malignant melanoma, the patient's prognosis would be favorable in countries with a higher intensity of solar radiation compared to countries where it is lower. The reality is that the prospects for patients diagnosed in Australia are better than those who were diagnosed in the Uk. Both populations are genetically similar, because the Australian population is largely British in origin [102]. An extensive retrospective study of data from 1993 to 2003 comparing the 5-year survival of patients in Yorkshire, in the Uk, (n=4170) and New south Wales, in Australia, (n=30 520) showed a relatively lower risk of death in New south Wales. This was attributed in particular to the more frequent occurrence of thinner tumor types (tumors with thickness < or = 1 mm) according to the Breslow classification in this area, which is due to the detection of tumors at an earlier stage. At the same time, Australian patients had higher average values of vitamin D [103]. Analysis of patient data purely from the United kingdom has shown that higher serum concentrations of 25-hydroxyvitamin D at the time of diagnosis are more frequent in thinner tumor types. The conclusions of these two studies was that the difference between these countries was based on different tumor thickness according to Breslow at the time of diagnosis, and the thickness was associated with the serum level of vitamin D [104]. Comparing two groups with a similar genetic background but a different environment is still the subject of ongoing studies looking to elucidate the circumstances of malignant melanoma. It is not yet clear whether these findings reflect the benefits of continuous exposure to solar radiation and connected to the use of vitamin D or a different pathogenesis of melanomas which occurs in people who are at increased risk of actinic skin damage [105].
Assessment of Vitamin D Status
Due to the presence of several forms of vitamin D and their different binding strengths to vDBp, vitamin D is a difficult analyte to determine. so-called Total vitamin D (25-oHD2 + 25oHD3) in blood serum is currently recommended as the best indicator of vitamin D in the human body due to its long biological half-life of over 250 hours (2-3 weeks) [106]. Assessing the effects of vitamin D in skin tissue is based on the assumption that the serum level reflects its production in the skin [107, 108]. The proportion of vitamin D obtained from food increases in importance during the winter months. As indicated above, part of this production is released into systemic circulation, but a certain part is always dependent on reactions leading to the formation of the active form of vitamin D directly in skin tissue. Based on this relationship, it is thought that the serum level of vitamin D is predominantly a reflection of its formation in the skin in places exposed to Uv light, and thus the local skin production of the active form of vitamin D.
As for patients with a confirmed diagnosis of MM or at a higher risk for this disease, to date no optimal serum level of vitamin D has yet been definitively determined [109]. Field and Newton-Bishop (2011) proposed a value between 70 and 100 nmol/L (28-40 ng/mL) in patients with a pre-established diagnosis, because studies show that higher serum levels of Vitamin D may in general affect tumor cell proliferation [110]. In a prospective cohort study performed in 872 patients, higher serum levels of Vitamin D at the time of diagnosis were associated with a lower value of the Breslow thickness classification (penetration depth measured in millimeters) of malignant melanoma [104]. There were further results which showed a lower risk of relapse in MM patients supplemented with Vitamin D compared to healthy controls [111]. Moreover, the progression of the disease is associated with a statistically significant reduction in serum levels of Vitamin D [79].
Vitamin D therapy in malignant melanoma - a potential adjuvant therapy
Until recently, recommended doses of Vitamin D were related to its effects on bones and the metabolism of calcium and phosphorus. Currently, based on meta-analyses, suitable doses and values are also being established for patients suffering from or at risk of cancer (Table 2) [105].
The theoretical treatment options of MM with Vitamin D include the induction of its formation by UV irradiation, a topical application and oral administration in the form of supplements. The specific food sources of Vitamin D mean that a completely alimentary form of intake can never meet daily needs [112]. In the first option, the cutaneous synthesis of the active form of Vitamin D occurs, and its effects in this case are the most comprehensive. The optimal period of exposure to UV radiation, which would minimize the damage to the DNA and would mean maximum benefit in the form of synthesis of Vitamin D, remains an unanswered question. Accordingly, peroral administration is still considered the safest option, but possibly undesirable systemic effects can be expected. What still remains unclear is whether the beneficial effects of Vitamin D produced in the skin are larger than those that follow taking an equivalent amount of Vitamin D in the form of supplements. Analogues of Vitamin D with a short side chain or completely lacking one are characterized by a lower hypercalcemic activity [113, 114, 115]. These analogues proved to be more effective at inhibiting the cell proliferation of malignant melanoma compared to normal melanocytes and keratinocytes [116].
Based on recent research work, adjuvant treatment with Vitamin D is suggested in the III. and IV. stages of malignant melanoma together with any oncological treatment. This is taken from evidence showing a higher incidence of advanced disease stages and disease progression in patients with Vitamin D deficiency. In stages I and II of the disease the prophylactic use of very high doses of Vitamin D (50,000 to 100,000 IU per day) are recommended if the serum level of Vitamin D is low (< 30 ng/ml).
Table 2. Serum concentration of Vitamin D and health effects
After reaching a serum level between 50-100 ng/ml, the dose is reduced to 4000-6000 IU, while serum calcium levels are simultaneously monitored. This approach could be particularly beneficial for patients at high risk of metastasis (mitotic active melanomas in the vertical growth phase, which are relatively thick, or with positive sentinel lymph node) [117].
Conclusion
Currently, Vitamin D is regarded as a fat-soluble steroid hormone whose genomic effects are mediated through binding to a specific receptor. Non-classical, pleiotropic effects are now attributed to Vitamin D alongside its confirmed, classical effects in relation to the preservation of healthy bones.
These non-classical effects are currently being observed in
autoimmune diseases [118],
cardiovascular diseases [119],
hypersensitivity to infections [120], the
development of cancer [121], but also over the course of
physiological aging [122].
Their role in skin cancer and notably in malignant melanoma is much more controversial. The reason is mainly in the skin’s ability to generate the active form of Vitamin D (1,25(OH)2D3) via keratinocytes and its local paracrine effects on the adjacent cells and therefore melanocytes and malignant melanoma cells.
Epidemiological studies have repeatedly drawn attention to the relationship between exposure to sunlight and the risk of malignant melanoma, but the nature of this relationship appears to be complex. The classical relationship of dose and risk - the higher the dose, the higher the risk - does not apply. The dominant risk factor is sunburn and not cumulative sun exposure. The process of sunburn likely leads to the suppression of immune reactions, which is subsequently involved in the course of malignant melanoma carcinogenesis. The hypothesis is that intense sun exposure induces both genetic changes, resulting in tumour antigenicity, and the inability of the immune system to detect those changes [123].
Epidemiological researchers are increasingly coming to a consensus on the role of Vitamin D in the prevention of many types of cancer, the most discussed being prostate, breast and colon cancer [124]. It is believed that Vitamin D itself is also able to influence the process of carcinogenesis in malignant melanoma, and this will probably include invasion and the formation of early metastasis. Higher serum levels of Vitamin D have been reported in patients with thinner types of MM, and their findings at the time of diagnosis were associated with a better prognosis and a lower risk of relapse.
However, the anti-cancer effects of vitamin D in this disease may be limited by the different mechanisms of resistance of tumor cells to the effects of vitamin D (reduction in the bioavailability of vitamin D, suppression of vDR expression and [13] changes in the expression of vDR coregulators) [125]. Investigations of these mechanisms will probably further extend the treatment and prevention options of malignant melanoma as [14] well as other malignancies.