UVA causes skin cancer, perhaps UVB (Vitamin D) prevents skin cancer
Vitamin D status and risk for malignant cutaneous melanoma: recent advances.
Eur J Cancer Prev. 2017 Jan 25. doi: 10.1097/CEJ.0000000000000334. [Epub ahead of print]
 ::Click on chart for details::
* Overview UV and vitamin D observes the following
* Controversy: UVA and/or UVB cause skin cancer
* Consensus: Vitamin D prevents and treats many cancers
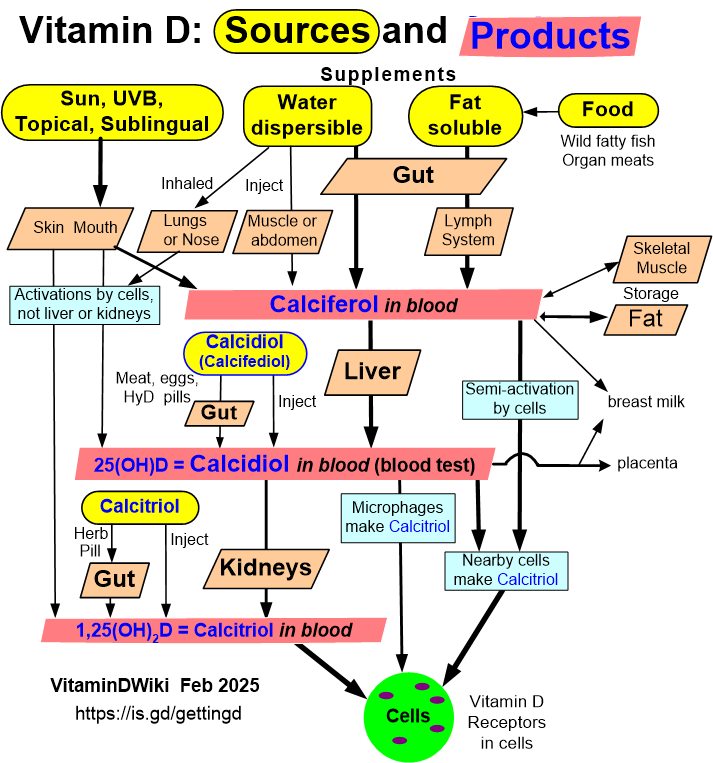
Vitamin D Receptor category has the following
{include}
Just the Cancer risks increased by VDR (as of Jan 2017)
||Risk increase|Health Problem
4.6|Breast Cancer
2 |Melanoma||__
::Click on chart for details::
* Overview UV and vitamin D observes the following
* Controversy: UVA and/or UVB cause skin cancer
* Consensus: Vitamin D prevents and treats many cancers
Vitamin D Receptor category has the following
{include}
Just the Cancer risks increased by VDR (as of Jan 2017)
||Risk increase|Health Problem
4.6|Breast Cancer
2 |Melanoma||__
📄 Download the PDF from VitaminDWiki
Figure 2
Ombra MN1, Paliogiannis P, Doneddu V, Sini MC, Colombino M, Rozzo C, Stanganelli I, Tanda F, Cossu A, Palmieri G.
Cutaneous malignant melanoma, whose incidence is increasing steadily worldwide, is the result of complex interactions between individual genetic factors and environmental risk factors. Ultraviolet radiation represents the most important environmental risk factor for the development of skin cancers, including melanoma. Sun exposure and early sunburn during childhood are the principal causes of cutaneous melanoma insurgence in adults, with double the risk relative to a nonexposed population. Consequently, ultraviolet protection has long been recognized as an important measure to prevent such a malignancy. Biological and epidemiological data suggest that vitamin D status could affect the risk of cancer and play a role in cancer prevention by exerting antiproliferative effects. Solar radiations are critical for vitamin D synthesis in humans; however, uncontrolled and intensive sun exposure is dangerous to skin health and may contribute toward the development of cutaneous malignant melanoma. An optimum balance between sun protection and exposure is thus advocated. Additional research is required to confirm the preventive role of vitamin D in melanoma incidence or a positive influence on patient outcome.
PMID: 28125434 DOI: 10.1097/CEJ.0000000000000334
Introduction
Cutaneous malignant melanoma (CMM) is one of the most frequent and aggressive forms of skin cancer that occurs in all age groups (Cossu et al., 2016). CMM arises from epidermal melanocytes, the cells responsible for the production of the melanin pigment. Risk factors associated with the development of malignant melanoma (MM) are multifactorial, with both genetic and environmental factors playing a role in its pathogenesis (Garbe and Leiter, 2009; Cossu et al., 2016). Epidemiological studies have identified risk factors including altered regulation of susceptibility genes (e.g. CDKN2A, CDK4, MC1R, ATM, and MX2), a family history of melanoma, hair and skin color, and ultraviolet (UV) light exposure (Holick, 2003; Nejentsev et al., 2004; Uitterlinden et al., 2004; Deeb et al., 2007; Casula et al., 2009; Welsh, 2011; Colombino et al, 2013, 2014; Pena-Chilet et al., 2013). Repeated sun exposure during childhood or adolescence resulting in sunburn is associated with an increased risk for melanoma; however, CMM can also arise in nonsun- exposed areas, also with different outcomes (Garbe and Leiter, 2009). Its incidence has increased markedly worldwide; in particular, the incidence of melanoma continues to increase in White populations irrespective of attempts to improve sun protection, underlining the need for further preventive measures and treatments (Linos et al., 2009; Rigel, 2010).
In recent years, attention has been focused on the possible role of vitamin D in cancer risk reduction and, in particular, in melanoma risk. Some studies suggest a protective role of vitamin D in melanoma, whereas results on the relationship between dietary intake of vitamin D and risk are controversial and there is inadequate evidence to suggest that vitamin D supplementation decreases the risk for melanoma. The relationship between vitamin D and melanoma seems to be more intricate compared with other cancers (Uitterlinden et al., 2004). In nonsun-exposed melanomas, systemic immunosuppression and other physiological aspects may influence the insurgence of the disease. A decrease in the serum levels of vitamin D may play a critical role in these patients and it has been reported that higher circulating levels of vitamin D are associated with a better prognosis in melanoma patients (Ahn et al., 2010; Zhao et al., 2014). The aim of this review is to summarize the most recent advances on the relationship between vitamin D and melanoma.
Vitamin D biosynthesis
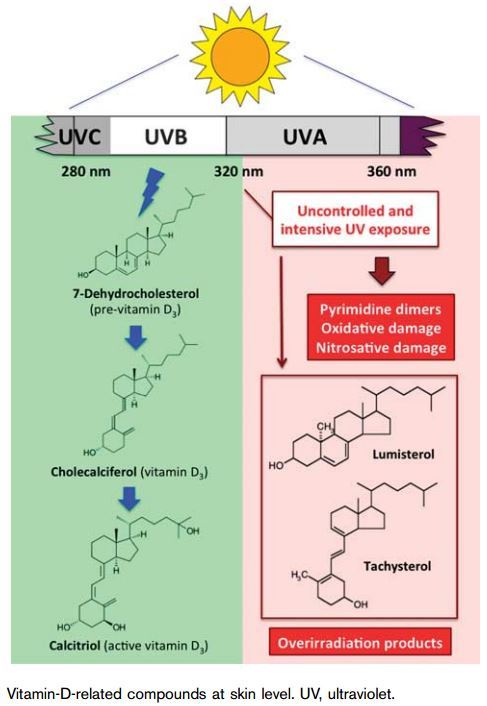
Vitamin D is a fat-soluble micronutrient, which plays a role in the maintenance of calcium and phosphate homeostasis, predominantly increasing the gut absorption of calcium and phosphate. Vitamin D is derived from two sources: endogenous (about 90% from synthesis in the skin - vitamin D3) and exogenous [dietary (Table 1 and Fig. 1) or supplements - vitamins D2 and D3 for the remaining 10%]. The substrate, 7-dehydrocholesterol (7-DHC), the penultimate compound in the cholesterol synthesis pathway, accumulates in the epidermis (Deeb et al., 2007; Pena-Chilet et al., 2013). UVB radiation to the skin transforms 7-DHC into previtamin D3, which undergoes nonenzymatic isomerization to form vitamin D3 (Fig. 2). Vitamin D3 is transferred into the blood stream by the vitamin D-binding protein, an a-globulin that has a high affinity for vitamin D and its metabolites.
To be physiologically active, vitamin D (either D2 or D3) must first be hydroxylated into 25-hydroxyvitamin D (25-OHD), predominantly in the liver (Figs 2 and 3) (Spina et al., 2006). Then, 25-OHD is subsequently hydroxylated into 1a,25-dihydroxyvitamin D [1,25 (OH)2D3] or calcitriol (active form of vitamin D), predominantly in the kidneys, although many tissues (including the skin) possess the capacity to produce 1a,25-OHD (Figs 2 and 3) (Spina et al., 2006). In fact, the entire metabolic pathway can be performed in UVB- irradiated skin and 1,25(OH)2D3 can be produced within 16 h. Figure 2 shows the biosynthesis of vitamin D in detail. Briefly, vitamin D is mainly synthesized in sun- exposed skin, where UV radiation B (280-320 nm) induces the photolysis of the B ring in 7-DHC. Starting from the 7-DHC substrate, a cascade of compounds is sequentially generated: previtamin D3 to vitamin D3, which in turn is transformed into 25-OHD, and, finally, 1,25(OH)2D3. Skin phenotypes and the UVB dose influence vitamin D levels (Parkin, 2011).
Table 1 Food sources of vitamin D as international units per 100 g
Biological activity
The primary role of vitamin D involves the regulation of bone metabolism and calcium-phosphorus homeostasis. However, over the past two decades, numerous in-vitro and in-vivo studies have evidenced several ‘noncalcemic’ or ‘extraskeletal’ effects of vitamin D (Holick, 2005). Reduced levels of vitamin D influence the onset and progression of several diseases such as autoimmune diseases, respiratory infections, diabetes mellitus type 1 and 2, hypertension and cardiovascular diseases, neuromuscular disorders, and cancer (Holick and Chen, 2008). The effects of vitamin D are largely correlated with the presence and activity of the nuclear vitamin D receptor (VDR). In fact, to exert its biological functions, 1,25 (OH)2D3 binds to the VDR and heterodimerizes with the retinoid X receptor (Spina et al., 2006). The hetero- dimeric complex interacts in the nucleus with specific sequences in the promoter region of vitamin D-responsive genes, known as the vitamin D-responsive element (VDRE) (Spina et al., 2006). It regulates the expression of more than 900 genes, among which are genes of cell cycle progression, differentiation, and apoptosis (Valverde et al., 1995; Bonilla et al., 2005; Graf et al., 2005; Lamason et al., 2005). These biological activities suggest that high levels of vitamin D metabolites may be protective against cancer (Deeb et al., 2007).
Calcitriol exerts significant antitumoral activity in vitro and in vivo in murine squamous cell carcinoma (SCC), rat metastatic prostatic adenocarcinoma Dunning (MLL) model systems, human prostatic adenocarcinoma (PC-3 and LNCaP), human breast, colon, and pancreatic cancer, as well as in leukemia, myeloma, and lymphoma lines (Holick, 2004). In fact, calcitriol regulates multiple signaling pathways involved in proliferation, apoptosis, differentiation, and angiogenesis, and it therefore has the potential to influence cancer development and growth (Holick, 2007; Weinstein et al., 2015).
Numerous effects of 1,25(OH)2D3 also include improvement of DNA repair processes, defense against reactive oxygen species (ROS), and immunomodulation (Rass and Reichrath, 2008; Reichrath and Reichrath, 2012). Despite its antiproliferative properties, the use of 1,25(OH)2D3 as a therapeutic agent is limited because of its hypercalcemic effects. However, the reduction or elimination of the cholesterol-type side chain significantly reduces or abolishes the calcemic effects; furthermore, a certain number of analogs of vitamin D3 have been synthesized, with the clinical activity of many of these still being investigated, both as single agents and in combination with other drugs (Rochel et al., 2001).
Fig. 1
As VDR is present in most cells in the body and calcitriol regulates 3-5% of the human genome, vitamin D activity is extensive, and can alter the defenses of the human body and most likely limit the progression of multiple diseases, including cancer (Fig. 3) (Stumpf et al., 1979; Spina et al., 2006). Among the many genes that are induced by calcitriol, CYP24A1 (also known as 24-hydroxylase) is particularly important; it encodes the enzyme that catalyzes the degradation of both 1,25 (OH)2D3 (calcitriol) and 25(OH)D3 (Prosser and Jones, 2004). Thus, the vitamin D activity is self-regulated as it may induce its own inactivation.
Vitamin D levels
UV radiation is the leading environmental factor involved in the development of skin cancer as well as melanoma. The synthesis of melanin in the skin represents a natural protective mechanism against UV-induced damage and carcinogenesis, but also limits vitamin D3 synthesis. Vitamin D deficiency is a worldwide problem, with insufficient vitamin D levels increasing the risk of developing obesity, diabetes, asthma, autoimmune disorders, infectious diseases, and some cancers. Several meta-analyses support the evidence that low 25-hydro- xyvitamin D3 serum level is a risk factor for many chronic diseases including cancer (Guerrieri-Gonzaga and Gandini, 2013). The minimum desirable serum level of hydroxyvitamin D3 has been suggested to be between 20 and 30 ng/ml (Norman et al., 2007). In 2011, the Institute of Medicine committee (Washington, DC, USA) reported guidelines stating that a level of 25-hydroxyvitamin D3 higher than 20 ng/ml is needed for good bone and general health for almost all individuals (Ross et al., 2011). However, the Institute of Medicine report did not consider clinical and demographic variables (race/ethnicity, body composition, sun exposure).
Recently, a meta-analysis from eleven prospective cohort studies suggests that the optimal 25-hydroxyvitamin D3 concentrations of between 30 and 40 ng/ml (75 and 100 nmol/l) are needed to reduce mortality (Zittermann et al., 2012). Nevertheless, the role of vitamin D in skin cancer risk is difficult to evaluate as serum levels of 25-OHD, as well as vitamin D intakes, are widely different from country to country from North to South Europe. Assays used to measure serum 25(OH)D3 levels are not standardized and can yield variable results. This needs to be taken into consideration when interpreting the relationship between vitamin D and disease.
An important issue is how much exposure to sunlight is required to produce sufficient levels of circulating 25- (OH)D3 for good health and whether this can be achieved without the deleterious effects of excessive sunlight exposure (such as skin cancer) or whether nutritional supplementation is needed. Several variables related to each individual (genetic constitution, skin type, area of sun-exposed skin, clothing, behavior) and environmental factors that influence the intensity and spectral range of UVB (such as latitude, season, and ozone layer) may be influential (Spina et al., 2006). Guidelines have been developed for personal sunlight exposure to attain desirable levels of vitamin D (Holick et al., 2011). 25-OHD is the major circulating form of vitamin D and, as a result, is used to determine the vitamin D status of both children and adults. The Endocrine Society’s practice guidelines recommend that children 1 year and older receive 600-1000IU daily and adults receive 1500-2000 IU daily, with the recommendation that obese individuals require 2-3 times more (Holick et al., 2011).
Sun exposure and melanoma
Melanoma risk is associated with the skin color types (I, II, and III in north Europe and types III, IV in central Europe populations) and with the gradient of annual UVB-UVA radiation. Scandinavia is located at latitudes (>54°N) where the annual UVB (280-315 nm) exposures are moderate and only of the order of 25% of the equatorial UVB exposures (Moan and Dahlback, 1995). Despite this, CMM is a significant health problem in Scandinavia. In 2008, the estimated age-adjusted incidence rate of CMM for women in Norway was 16.5, compared with 8.9 in France, 5.6 in Spain, 8.7 in Italy, and 12.6 in Germany (Ferlay et al., 2010).
The relationship between annual exposure to solar radiation (weighted using the CIE erythema action spectrum) and incidence rates of basal cell carcinoma (BCC), SCC, and CMM for Scandinavia, England, New Zealand, and Australia analyzed using logarithmic functions is similar with respect to skin types (Moan et al., 2008). For SCC and BCC, the risk of disease is similar for men and women, whereas for CMM, the incidence rate is significantly higher for men than for women. The reason for this sex difference is likely to be that women in north Europe, New Zealand, and Australia tend to expose themselves intermittently to high doses during summer vacations compared with the men, who have lower exposure patterns (Moan et al., 1990, 2013).
Individuals with dark, African skin (types V-VI) have a 20-fold lower risk of developing skin cancers than White individuals living at the same latitude (Wu et al., 2011). Individuals of color living near the equator and exposed to sunlight on a daily basis have blood levels of 25(OH)D of 40-60 ng/ml. Their skin was designed to produce an adequate amount of vitamin and the melanin pigmentation prevents the damaging effects, minimizing the risk of nonmelanoma skin cancer (NMSC). Asians have an intermediate risk (Wu et al., 2011). A recent investigation shows that the CMM mortality decreases with decreasing latitude in Europe as a whole (Spina et al., 2006; Shipman et al., 2011; Moan et al, 2013).
In summary, the evidence that melanoma is caused by sun exposure comes from epidemiological data showing that melanoma incidence is highest where fair-skinned individuals (susceptible to sunburn) live at low latitudes. Furthermore, among white-skinned individuals, the risk is highest for those who report sunburn and sun exposure on holidays. The relationship between sun exposure and melanoma risk is, however, complex. Overall, the genetic-epidemiological data suggest that intermittent sun exposure and sunburn are factors that are responsible for the marked increase in melanoma incidence.
Factors influencing vitamin D production
During exposure to sunlight, the UVB radiation enters the skin and activates the reactions for the production of previtamin D3 as we reported before. The UVB photons also signal melanocytes to increase the synthesis of melanin. Melanin acts as a natural sunscreen and is efficiently packaged into melanosomes that migrate upward to the upper layers of the epidermis, where they efficiently absorb UVB and UVA radiation. An increase in skin pigmentation is inversely related to the number of UVB photons that can penetrate into the epidermis and dermis. Thus, the efficiency in utilizing UVB photons to produce vitamin D3 in the skin is inversely related to the amount of skin pigmentation. An individual with deep skin pigmentation of African origin (skin type 5), who is exposed to the same amount of sunlight as an individual with minimum skin pigmentation of Celtic or Scandinavian origin (skin type 2), will produce less vitamin D3 compared with that produced in the lighterskinned individuals (Weinstein et al., 2015). Aging causes a decrease in the amount of 7-DHC in the epidermis; elders exposed to the same amount of sunlight as a young adult will produce a lower amount of vitamin D compared with a young adult (Weinstein et al., 2015).
The angle at which sunlight penetrates the Earth’s atmosphere also influences the production of vitamin D in the skin. This angle, known as the zenith angle, is related to season, time of day, and latitude. At the lowest zenith angle, more UVB photons penetrate the Earth’s surface. During the winter (i.e. November-February) above and below 35° latitude, the zenith angle is so oblique that UVB photons are absorbed by the stratospheric ozone layer. As a result, very little vitamin D can be produced in human skin. At very high latitudes, such as in Norway and Canada, little vitamin D is produced between the months of October and March. Latitude, season, and time of day influence the production of vitamin D in the skin (Weinstein et al., 2015).
To solve the problem of these variables associated with sun-induced vitamin D synthesis including time of day, cloud cover, season, latitude, and skin type, a free app has been developed that provides the user not only with useful information for how much vitamin D is being produced but also alerts when they are at risk for overexposure to sunlight. Under some circumstances, dietary supplementation may be necessary and perhaps preferable to sunlight.
Vitamin D and cancer
Epidemiological studies have reported that individuals who live in higher latitudes are at a higher risk of developing breast, colon, ovarian, and prostate cancers; indeed, mortality rates in both men and women have been related to their exposure to sunlight (Grant, 2002). Several reports indicated that some of the most common cancers were significantly reduced in individuals with higher circulating levels of the antiproliferative hormone 25(OH)D, and that adequate vitamin D supplementation and sensible sunlight exposure are among the front-line factors of prophylaxis for a wide spectrum of tumors (Pludowski et al., 2013). Indeed, the increased levels of 25(OH)D levels were found to prevent some cancers through downregulation of cell growth (Stumpf et al., 1979; Spina et al., 2006). However, it was difficult to understand how the increased exposure to sunlight could aid in decreasing the risk of common cancers as it is known that any significant increase in vitamin D intake or exposure to sunlight does not increase blood levels of 1,25(OH)D. Some clues to the pathological mechanisms were provided by observations that normal and tumor prostate cells may express a functional 1-OHase enzyme similar to that detected at the cutaneous level (Schwartz et al., 1998). Since the initial observation, it is now recognized that normal and tumor cells in colon, breast, and other tissue types possess the enzymatic machinery to convert 25(OH)D directly into 1,25(OH)2D (Spina et al., 2006). Thus, it seems that adequate levels of 25- (OH)D, probably above 30 ng/ml, considering that the demarcations between deficiency (< 20 ng/ml), insufficiency (20-30 ng/ml), and optimal (30-80 ng/ml) serum concentrations are controversial, may become the substrate for the extrarenal 1-OHase in several tissues (Haines and Park, 2012). The production of 1,25(OH)2D at multiple locations may be necessary to maintain and regulate genes involved in controlling cell growth and survival as well as to prevent cells from a carcinogenic transformation.
Observational studies on vitamin D levels and melanoma
Results on the relationship between vitamin D level and melanoma risk are controversial. Few studies have prospectively measured serum vitamin D before the development of melanoma, whereas most investigations have assessed serum vitamin D close to the time of diagnosis. In additional studies, vitamin D status at the time of diagnosis has been evaluated and patients were followed for survival. Moreover, the correlations between serum vitamin D levels and melanoma risk or serum vitamin D levels and survival of melanoma patients should be distinctly taken into consideration.
For a number of analyses, there was no evidence of increased melanoma risk associated with serum vitamin D. Epidemiological studies designed to look at the risk associated with vitamin D levels in the serum have many limitations including the fact that the levels likely change over time so that a single measure does not necessarily represent the complete picture. It is also not clear as to what is the most relevant measure (average vitamin D over a period or the lowest level of vitamin D) to indicate the most critical period. The risk could also be modified by the individual’s ability to utilize the circulating vitamin D. Major et al. (2012) carried out the first prospective study evaluating the relationship between prediagnostic serum vitamin D concentrations and subsequent melanoma risk. The results of this study indicate no statistically significant association between serum 25-(OH)D levels and melanoma, although there is a suggested protective association in the second quartiles compared with the lowest levels (Major et al., 2012). In other studies, deeper Breslow thickness or higher stage was associated with lower serum vitamin D or both (Newton-Bishop et al., 2009; Nurnberg et al., 2009; Gambichler et al., 2012). Newton-Bishop et al., (2009) showed that serum 25-hydroxyvitamin D3 levels were inversely correlated with tumor thickness among 941 patients with melanoma, although there was no difference in serum vitamin D levels between healthy controls (n = 114) and the patients. Another study showed no direct relationship between high or low levels of vitamin D and the occurrence and severity of melanoma (Garland et al., 2003). In contrast, Nurnberg et al. (2009) evidenced that among patients with MM, significantly reduced serum 25(OH)D levels were found in stage IV patients compared with stage I patients, and those with low 25-(OH)D serum levels (< 10 ng/ml) probably develop earlier distant metastatic disease compared with those with higher 25 (OH)D serum levels (>20 ng/ml).
Data on the relationship between dietary intake of vitamin D and melanoma risk are also uncertain. Tang et al. (2011) found that daily supplementation with 1000 mg of calcium and 400 IU of vitamin D did not reduce the overall incidence of NMSC or melanoma in a large randomized double-blinded placebo-controlled trial. Vinceti et al. (2011) examined the association between vitamin D and melanoma risk through a population-based case-control study. They described an inverse association between dietary intake of vitamin D and melanoma risk, in particular among men and older patients. Finally, there is evidence that patients with CMM who strictly avoid sun exposure might benefit from 25-OHD supplements that are sufficient to maintain serum levels above 30 ng/ml (Gambichler et al., 2012; Zittermann et al., 2012). In a recent study, Fang et al. (2016) simultaneously examined vitamin D and C-reactive protein levels in a hospital-based cohort of patients with melanoma in relation to demographics, tumor histopathology, disease stage, and clinical outcome measures. They found that a lower level of vitamin D was associated with higher C-reactive protein, fall/winter months of blood draw, thicker and more ulcerated primary tumors, and advanced melanoma stage; they also found that a lower level of vitamin D was associated independently with poorer overall survival and disease-free survival (Fang et al., 2016). More research is needed to determine whether 25(OH)D levels play a role in tumor control and progression of CMM.
As epidemiological studies have reported uncertain results and given the difficulties in assessing the risk of melanoma in response to serum levels of vitamin D, investigations have been directed toward the study of the vitamin D pathway, and in particular on the VDR receptor and its expression in melanocytes. Therefore, in recent studies, polymorphisms of VDR genes (which are often overexpressed in melanoma cells) are associated with the occurrence of several cancers including melanoma (Maruyama et al., 2006; Hou et al., 2015).
Vitamin D receptor
The most studied gene in the vitamin D pathway is the VDR gene located on chromosome 12q13.11. The gene has 11 exons, with more than 600 single-nucleotide polymorphisms (SNPs) having been identified within its coding region. This receptor is a nuclear transcription factor that belongs to the steroid hormone superfamily of receptors.
Nuclear receptors integrate hormonal, dietary, and other extracellular signals into cell fate decisions by regulation of gene expression and repression of a host of common gene targets. The VDR mediates the major cellular effects of vitamin D. Active hormone 1,25(OH)3D3, by binding to VDR, regulates various signaling pathways involved in cell cycle progression, differentiation, and apoptosis, also implicated in cancer development and progression (Welsh, 2011).
Many epidemiological studies have evaluated associations between VDR variants and various types of cancer including those of the breast, colorectal region, ovary, and prostate (Maruyama et al., 2006). Similarly, the associations of VDR polymorphisms with skin cancer risk are known. In a mouse model, dysfunctional VDR increased susceptibility to skin cancer following exposure to 7,12-dimethylbenz[a]anthracene or UV light (Pena- Chilet et al., 2013). Given the genetic and environmental interactions between VDR and UV light exposure during the development of skin cancer, many epidemiological studies have also examined associations between VDR variants and melanoma risk. However, the results of these studies have not been consistent. Only a few SNPs, considered functional, occur at a high frequency in White populations and may influence receptor affinity, binding to nuclear DNA, RNA transcription, and protein synthesis (Uitterlinden et al., 2004).
A recent study of VDR polymorphisms including Fok1, Bsm1, Apal, and Taq1 in a White population was carried out to define roles of the polymorphisms in skin cancer risk, incidence, and development. The results of this study show that Fok1 polymorphism is associated with an overall significantly increased risk of skin cancer (Zhao et al., 2014). The Taql polymorphism could contribute toward NMSC susceptibility, whereas the Apal polymorphism is associated with the development of skin cancer. No significant association was observed between the Bsml polymorphism and the risk of skin cancer. A meta-analysis study shows that Fokl, Taql, and Apal may be the susceptibility biomarkers for skin cancer in Whites (Zhao et al., 2014).
A trend of cancer risk reduction with the VDR BsmI B allele was observed for many cancer sites. In particular, a meta-analysis was carried out to investigate the role of the VDR BsmI polymorphism in cancer risk, even according to different ethnicities (Raimondi et al., 2014). The metaanalysis included 73 studies with 45 218 cases and 52 057 controls. A significant 6-7% reduction of cancer risk at any site was found, respectively, for carriers of Bb and for carriers of the BsmI BB genotype compared with bb carriers. For skin cancer, a significant risk reduction was observed for Bb carriers (summary odds ratio = 0.86; 95% confidence interval = 0.76-0.98). When the analysis was stratified by ethnicity, a significantly decreased risk for both Bb and BB compared with the bb genotype among Whites was still observed for any cancer site. Among other ethnic groups, the inverse association was still present, but did not reach statistical significance. In conclusion, a weak effect of the BsmI B allele in reducing cancer risk at any site, especially of the skin, was reported (Raimondi et al., 2014).
VDR variants have been found to be associated with the risk of developing melanoma. In fact, in a recent epidemiological meta-analysis study, 10 eligible studies were identified and six VDR variants were evaluated (Apal, Bsml, Cdx2, EcoRV, Fokl, and Taql) in a total of 4961 melanoma patients and 4605 controls (Hou et al., 2015). It has been reported that the VDR variants Fokl and Bsml may influence the susceptibility to developing melanoma, but not the other four variants Apal, Cdx2, EcorV, and Taeql. The dominant genetic model suggested that the allele f carriers showed an 18% (pooled odds ratio = 1.18, 95% confidence interval = 1.07-1.29; I2 = 0%) increased risk for melanoma compared with homozygote FF, whereas for BsmI, under the dominant genetic model, a 15% decrease of melanoma risk was found for those with the BB or the Bb genotype compared with those with the bb genotype. The VDR variants Fokl and Bsml may influence the susceptibility to the development of melanoma, although further studies are required to confirm these conclusions definitively (Hou et al., 2015).
Genes involved in the vitamin D pathway
In addition to VDR, a key protein in vitamin D metabolism is the vitamin D-binding protein (DBP), and the gene encoding the DBP (GC, group-specific component) plays an important role in the vitamin D pathway. Recent studies using genome-wide association (GWAS) report that an SNP in the GC gene is associated with serum levels of vitamin D3 (Wang et al., 2010). Additional genes related to the production of the active form of vitamin D are probably involved in vitamin D status (Ahn et al, 2010). In a previous GWAS of 4501 individuals of European ancestry, SNPs were identified in the gene encoding group-specific component (vitamin D binding) protein, GC, on chromosome 4q12-13, and suggestive signals for SNPs in or near three other genes involved in vitamin D synthesis or activation (Ahn et al., 2010). Rs3829251 is on chromosome 11q13.4 in NADSYN1, encoding nicotinamide adenine dinucleotide synthetase, which was in high linkage disequilibrium with rs1790349, located in DHCR7, the gene encoding 7-DHC reductase that synthesizes cholesterol from 7-DHC. rs6599638, in the region with open-reading frame 88 (Cl0orf88) on chromosome 10q26.13 near ACADSB (acyl-coenzyme A dehydrogenase), is involved in cholesterol and vitamin D synthesis, and rs2060793, on chromosome 11p15.2 in CYP2R1 (cytochrome P450, family 2, subfamily R, polypeptide 1), encodes a key C-25 hydroxylase that converts vitamin D3 into an active vitamin D receptor ligand (Deeb et al., 2007; Pena-Chilet et al., 2013). SNPs in these four regions in 2221 additional samples were associated with 25(OH)D through meta-analysis with the GWAS data (Ahn et al, 2010). In particular, SNPs for GC, NADSYN1/DHCR7, and CYP2R, but not for Cl0orf88, were associated with 25(OH)D concentrations (Ahn et al., 2010). In the GC gene, at least six nonsynonymous SNPs are described, two of them with common frequency (rs7041 and rs4588) in White populations. The association between GC variants (rs12512631) and vitamin D levels in plasma has already been tested and proved in various lung diseases, but few studies support the association between polymorphisms on GC and melanoma (Ahn et al., 2010; Pena-Chilet et al., 2013; Hou et al., 2015).
Ultimately, several studies investigated DBP serological levels and GC polymorphisms in association with cancer risk, with controversial results. Thus, a meta-analysis was carried out to investigate these associations and a total of 28 independent studies were included for the following tumors: BCC, bladder, breast, colon-rectum, endometrium, liver, esophagus, stomach, melanoma, pancreas, prostate, and kidney (Tagliabue et al., 2015). The GC polymorphisms rs2282679, rs12512631, rs7041, rs4588, rs17467825, rs1155563, and rs1352844 were considered. Only a borderline decrease was found in cancer risk for patients with high levels compared with low levels of DBP, suggesting a role of DBP in cancer etiology, which should be confirmed in further studies (Tagliabue et al, 2015).
VDR and the p53 gene
Depending on the cell type and context, both VDR signaling and p53 signaling regulate many cellular functions that are of relevance for cancer development, including proliferation, differentiation, apoptosis, and cell survival; consequently, vitamin D and p53-signaling pathways have a significant impact on spontaneous or carcinogen-induced malignant transformation of cells (Maruyama et al., 2006). Furthermore, the VDR and p53 genes represent two important tumor suppressors in several tissues (Evans et al., 1998; Palmer et al., 2004). Mutations in genes encoding proteins of the p53 pathway represent a hallmark of many if not all types of cancer.
Much of the tumor-suppressor function in the skin may be mediated through the interaction of the VDR and p53 pathways, either by mutual activation or by inhibition (Maruyama et al., 2006). UV radiation induces different forms of DNA lesions, which are generated either photochemically and directly or indirectly by UV activation of several photoreceptors, which have the capacity to modulate the cellular redox equilibrium, thereby generating ROS (Reichrath and Reichrath, 2012). Both UV- induced and ROS-induced damage activates p53. UV irradiation induces p53 to stimulate skin pigmentation by pro-opiomelanocortin derivatives including a-melano- cyte-stimulating hormone and adrenocorticotropic hormone, reducing the cutaneous synthesis of vitamin D (Gupta et al., 2007). Cancer cells often contain abundant mutant p53 (mutp53) protein, which may contribute actively toward tumor progression by a gain-of-function mechanism. The VDR response element was identified as over-represented in promoter sequences bound by mutp53. Stambolsky et al. (2010) reported that mutp53 can interact functionally and physically with VDR. Mutp53 is recruited into VDR-regulated genes, augmenting the transactivation of some genes or decreasing the expression of others. Furthermore, mutp53 increases the nuclear accumulation of VDR and converts vitamin D into an antiapoptotic agent. Thus, p53 status can influence the biological impact of vitamin D on tumor cells and should be examined when considering vitamin D analogs for cancer therapy.
In summary, the p53 tumor-suppressor gene, once mutated, not only loses its tumor-suppressor activity but gains oncogenic functions. Indeed, p53 has been considered an important target for cancer therapy. Similarly, vitamin D and its analogs are being evaluated as potential anticancer agents. An interaction between p53 and VDR has been reported, with clinical implications, suggesting that p53 status should be considered when contemplating vitamin D analogs for cancer therapy.
Genes influencing skin pigmentation and SLC45A2
In humans, only a few genes show effects on normal variations of skin pigmentation associated with UV radiation. The strongest evidence of pigmentation- mutation associations is found in the pigmentary genes MC1R, ASIP, SLC24A5, SLC45A2, TYR, OCA2, and KITLG (Bonilla et al., 2005; Graf et al., 2005; Lamason et al., 2005; Yuasa et al., 2007; Nan et al., 2009; de Gruijter et al., 2011). Among these, SLC45A2 serves a major function in the process of melanin synthesis by controlling the activity and traffic of tyrosinase to the melano- somes and maintaining the melanosomal pH (Lucotte et al., 2010; Dooley et al., 2013). SLC45A2, also known as MATP or AIM1, is a membrane-associated transporter gene located on chromosome 5p and consists of seven exons spanning a region of ~ 40 kb. Human mutations in this gene can cause oculocutaneous albinism type 4 (Newton et al., 2001).
The involvement of genetic variants of SLC45A2 in melanoma susceptibility has also been investigated. In fact, the variant 374L is protective against melanoma in different European populations (Dooley et al., 2013). An association of two common SNPs in SLC45A2, Leu374Phe (L374F, rs16891982), and Glu272Lys (E272K, rs26722) has been found with human pigmentation variations in European descendants from Australia (presumably of North European origin). The ancestral 374L allele, which is fixed in African populations, would contribute to an optimal eumelanin production, whereas the 374F allele, which is almost fixed in European populations, would give rise to an acidic melanosomal environment that negatively affects tyrosinase activity, hence leading to a lighter pigmentation (Lucotte et al., 2010).
The L374F SNP is significantly associated with melanoma, and the 374F (the ‘light’ pigmentation allele) represents a risk factor for melanoma (Cochran-Armitage trend test assuming an additive model, P: 4.36E — 06), although a nonuniform allele frequency is found across the geography according to the intensity of the incident UV radiation; this nonhomogeneous allele distribution suggests that positive selection has acted on the SLC45A2 region (Lopez et al., 2014).
Conclusion
In recent years, there has been growing interest in understanding the link between vitamin D status and melanoma as well as other cancers and chronic diseases. The relationship of serum levels and genetic factors of vitamin D with melanoma risk and melanoma mortality is actually not completely clear. There are few strong epidemiological studies to confirm the hypothesis that higher vitamin D levels might protect from melanoma, although a number of cohort studies have addressed a possible protective effect of vitamin D. In addition, there are insufficient indications to recommend vitamin D supplementation to decrease melanoma risk.
Certainly, vitamin D has clear antiproliferative activity on melanoma cell lines in vitro . There is evidence of reduced expression of the vitamin D receptor with progression from nevi through primary to metastatic melanoma. These observations suggest that if vitamin D is antiproliferative for melanoma cells in vivo, then those cells might be less likely to respond to the antiproliferative effects of vitamin D as progression occurs.
Other results indicate that high levels of vitamin D are correlated with the development of less aggressive tumors
Some studies report that normal levels of vitamin D3 at the time of diagnosis are associated with a better prognosis in patients with melanoma.
High circulating vitamin D concentration has been found to be associated with reduced melanoma progression and improved survival.
Furthermore, reduced vitamin D serum levels have been reported in patients with stage IV melanoma compared with those with stage I.
An unresolved question remains as to which of the many described biological effects of vitamin D might have a protective effect for melanoma patients and what might be the optimal blood level to achieve those putative beneficial effects. The in-vitro antiproliferative effect of vitamin D added to melanoma cell cultures is convincing, and the Leeds Melanoma Cohort data suggest that primaries in individuals included in the study were thinner in patients with higher 25-hydroxyvitamin D2/D3 levels, which is consistent with an antiproliferative effect. Vitamin D, moreover, may exert pleomorphic effects including those on new blood vessel formation and immunity, and some of these effects might actually be deleterious for cancer patients. The reported effects of vitamin D on the immune system are extremely complex. If vitamin D supplementation suppresses adaptive immunity, then that would be a potentially harmful effect for melanoma patients. High doses of vitamin D should also be avoided. The evidence that vitamin D levels might influence melanoma risk remains uncertain; however, it should also be pointed out that studies of sufficient size to address this issue have not been carried out. In addition, there is evidence that patients with MM who strictly avoid sun exposure might benefit from 25- (OH)D supplements that are sufficient to maintain serum levels above 30 ng/ml . Given the interest in using vitamin D to reduce cancer risk, more research is warranted to establish its role in the control and progression of melanoma and whether vitamin D supplements can reduce risk and progression and improve outcomes.
Although the association between vitamin D and melanoma risk is still the object of considerable discussion, the potential effect of vitamin D on the risk of melanoma merits accurate consideration.
References
Ahn J, Yu K, Stolzenberg-Solomon R, Simon KC, McCullough ML, Gallicchio L, et al. (2010). Genome-wide association study of circulating vitamin D levels. Hum Mol Genet 19:2739-2745.
Bonilla C, Boxill LA, Donald SA, Williams T, Sylvester N, Parra EJ, et al. (2005). The 8818 G allele of the agouti signaling protein (ASIP) gene is ancestral and is associated with darker skin color in African Americans. Hum Genet 116:402-406.
Casula M, Muggiano A, Cossu A, Budroni M, Caraco C, Ascierto PA, etal. (2009). Role of key-regulator genes in melanoma susceptibility and pathogenesis among patients from South Italy. BMC Cancer 9:352.
Colombino M, Lissia A, Franco R, Botti G, Ascierto PA, Manca A, et al. (2013). Unexpected distribution of cKIT and BRAF mutations among southern Italian patients with sinonasal melanoma. Dermatology 226:279-284.
Colombino M, Sini MC, Lissia A, De Giorgi V, Stanganelli I, Ayala F, et al. (2014). Discrepant alterations in main candidate genes among multiple primary melanomas. J Trans Med 12:11 7.
Cossu A, Casula M, Cesaraccio R, Lissia A, Colombino M, Sini MC, et al. (2016). Epidemiology and genetic susceptibility of malignant melanoma in North Sardinia, Italy. Eur J Cancer Preven [Epub ahead of print].
de Gruijter JM, Lao O, Vermeulen M, Xue Y, Woodwark C, Gillson CJ, et al. (2011). Contrasting signals of positive selection in genes involved in human skin-color variation from tests based on SNP scans and resequencing. Investig Genet 2:24.
Deeb KK, Trump DL, Johnson CS (2007). Vitamin D signaling pathways in cancer: potential for anticancer therapeutics. Nat Rev Cancer 7:684-700.
Dooley CM, Schwarz H, Mueller KP, Mongera A, Konantz M, Neuhauss SC, et al. (2013). Slc45a2 and V-ATPase are regulators of melanosomal pH homeostasis in zebrafish, providing a mechanism for human pigment evolution and disease. Pigment Cell Melanoma Res 26:205-21 7.
Evans SR, Nolla J, Hanfelt J, Shabahang M, Nauta RJ, Shchepotin IB (1998). Vitamin D receptor expression as a predictive marker of biological behavior in human colorectal cancer. Clin Cancer Res 4:1591-1595.
Fang S, Sui D, Wang Y, Liu H, Chiang YJ, Ross MI, et al. (2016). Association of vitamin D levels with outcome in patients with melanoma after adjustment for C-reactive protein. J Clin Oncol 34:1 741 -1747.
Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM (2010). Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer 127:2893-2917.
Gambichler T, Scola N, Bechara FG (2012). Significantly decreased methyl- thioadenosine phosphorylase expression in malignant melanoma. Am J Dermatopathol 34:777-779.
Garbe C, Leiter U (2009). Melanoma epidemilogy and trends. Clin Dermatol 27:3-9.
Garland CF, Garland FC, Gorham ED (2003). Epidemiologic evidence for different roles of ultraviolet A and B radiation in melanoma mortality rates. Ann Epidemiol 13:395-404.
Graf J, Hodgson R, van Daal A (2005). Single nucleotide polymorphisms in the MATP gene are associated with normal human pigmentation variation. Hum Mutat 25:278-284.
Grant WB (2002). An estimate of premature cancer mortality in the US due to inadequate doses of solar ultraviolet-B radiation. Cancer 94:1867-1875.
Guerrieri-Gonzaga A, Gandini S (2013). Vitamin D and overall mortality. Pigment Cell Melanoma Res 26:16-28.
Gupta R, Dixon KM, Deo SS, Holliday CJ, Slater M, Halliday GM, et al. (2007). Photoprotection by 1,25 dihydroxyvitamin D3 is associated with an increase in p53 and a decrease in nitric oxide products. J Invest Dermatol 127:707-715.
Haines ST, Park SK (2012). Vitamin D supplementation: what’s known, what to do, and what’s needed. Pharmacotherapy 32:354-382.
Holick MF (2003). Evolution and function of vitamin D. Recent Results Cancer Res 164:3-28.
Holick MF (2004). Vitamin D: importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am J Clin Nutr 79:362-371.
Holick MF (2005). Vitamin D: photobiology, metabolism, mechanism of action and clinical application. In: Favus MJ, editor. Primer on the metabolic bone diseases and disorders of mineral metabolism, 5th ed. Washington, DC: American Society for Bone and Mineral Research. pp. 129-136.
Holick MF (2007). Vitamin D deficiency. N Engl J Med 357:266-281.
Holick MF, Chen TC (2008). Vitamin D deficiency: a worldwide problem with health consequences. Am J Clin Nutr 87:S1080-S1086.
Holick MF, Binkley NC, Bischoff-Ferrari HA, Gordon CM, Hanley DA, Heaney RP, et al. (2011). Evaluation, treatment, and prevention of vitamin D deficiency: an Endocrine Society clinical practice guideline. J Clin Endocrinol Metab 96:1911-1930.
Hou W, Wan X, Fan J (2015). Variants Fok1 and Bsm1 on VDR are associated with the melanoma risk: evidence from the published epidemiological studies. BMC Genet 16:14.
Lamason RL, Mohideen MA, Mest JR, Wong AC, Norton HL, Aros MC, et al. (2005). SLC24A5, a putative cation exchanger, affects pigmentation in zeb- rafish and humans. Science 310:1782-1786.
Linos E, Swetter SM, Cockburn MG, Colditz GA, Clarke CA (2009). Increasing burden of melanoma in the United States. J Invest Dermatol 129:1666-1674.
Lopez S, Garcia O, Yurrebaso I, Flores C, Acosta-Herrera M, Chen H, et al. (2014). The interplay between natural selection and susceptibility to melanoma on allele 374F of SLC45A2 gene in a South European population. PLoS One 9:e104367.
Lucotte G, Mercier G, Dieterlen F, Yuasa I (2010). A decreasing gradient of 374F allele frequencies in the skin pigmentation gene SLC45A2, from the north of West Europe to North Africa. Biochem Genet 48:26-33.
Major JM, Kiruthu C, Weinstein SJ, Horst RL, Snyder K, Virtamo J (2012). Prediagnostic circulating vitamin D and risk of melanoma in men. PLoS One 7: e3511 2.
Maruyama R, Aoki F, Toyota M, Sasaki Y, Akashi H, Mita H, et al. (2006). Comparative genome analysis identifies the vitamin D receptor gene as a direct target of p53-mediated transcriptional activation. Cancer Res 66:4574-4583.
Moan J, Dahlback A (1995). Predictions of health consequences of changing UV- fluence. In: Dubertret L, Santus R, Morliere P, editors. Ozone, sun, cancer, 1st ed. Paris: Les Editions Inserm. pp. 87-100.
Moan J, Dahlback A, Setlow RB (1990). Epidemiological support for an hypothesis for melanoma induction indicating a role for UVA radiation. Photochem Photobiol 70:243-247.
Moan J, Porojnicu AC, Dahlback A (2008). Ultraviolet radiation and malignant melanoma. Adv Exp Med Biol 624:104-116.
Moan J, Grigalavicius M, Baturaite Z, Juzeniene A, Dahlback A (2013). North- South gradients of melanomas and non-melanomas: a role of vitamin D? Dermatoendocrinol 5:186-191.
Nan H, Kraft P, Hunter DJ, Han J (2009). Genetic variants in pigmentation genes, pigmentary phenotypes, and risk of skin cancer in Caucasians. Int J Cancer 125:909-917.
Nejentsev S, Godfrey L, Snook H, Rance H, Nutland S, Walker NM, et al. (2004). Comparative high resolution analysis of linkage disequilibrium and tag single nucleotide polymorphisms between populations in the vitamin D receptor gene. Hum Mol Genet 13:1633-1639.
Newton JM, Cohen-Barak O, Hagiwara N, Gardner JM, Davisson MT, King RA, et al. (2001). Mutations in the human orthologue of the mouse underwhite gene (uw) underlie a new form of oculocutaneous albinism, OCA4. Am J Hum Genet 69:981 -988.
Newton-Bishop JA, Beswick S, Randerson-Moor J, Chang YM, Affleck P, Elliott F, et al. (2009). Serum 25-hydroxyvitamin D3 levels are associated with Breslow thickness at presentation and survival from melanoma. J Clin Oncol 27:5439-5444.
Norman AW, Bouillon R, Whiting SJ, Vieth R, Lips P (2007). 13th Workshop consensus for vitamin D nutritional guidelines. J Steroid Biochem Mol Biol 103:204-205.
Nurnberg B, Graber S, Gartner B, Geisel J, Pfohler C, Schadendorf D, et al. (2009). Reduced serum 25-hydroxyvitamin D levels in stage IV melanoma patients. Anticancer Res 29:3669-3674.
Palmer HG, Larriba MJ, Garcia JM, Ordonez-Moran P, Pena C, Peiro S, et al. (2004). The transcription factor SNAIL represses vitamin D receptor expression and responsiveness in human colon cancer. Nat Med 10:917-919.
Parkin DM (2011). The fraction of cancer attributable to lifestyle and environmental factors in the UK in 2010. Br J Cancer 105:S2-S5.
Pena-Chilet M, Ibarrola-Villava M, Martin-Gonzalez M, Feito M, Gomez- Fernandez C, Planelles D, et al. (2013). rs12512631 on the group specific complement (Vitamin D-Binding Protein GC) implicated in melanoma susceptibility. PLoS One 8:e59607.
Pludowski P, Holick MF, Pilz S, Wagner CL, Hollis BW, Grant WB, et al. (2013). Vitamin D effects on musculoskeletal health, immunity, autoimmunity, cardiovascular disease, cancer, fertility, pregnancy, dementia and mortality - a review of recent evidence. Autoimmun Rev 12:976-989.
Prosser DE, Jones G (2004). Enzymes involved in the activation and inactivation of vitamin D. Trends Biochem Sci 29:664-673.
Raimondi S, Pasquali E, Gnagnarella P, Serrano D, Disalvatore D, Johansson HA, et al. (2014). BsmI polymorphism of vitamin D receptor gene and cancer risk: a comprehensive meta-analysis. Mutat Res 769:17-34.
Rass K, Reichrath J (2008). UV damage and DNA repair in malignant melanoma and nonmelanoma skin cancer. Adv Exp Med Biol 624:162-178.
Reichrath J, Reichrath S (2012). Hope and challenge: the importance of ultraviolet (UV) radiation for cutaneous vitamin D synthesis and skin cancer. Scand J Clin Lab Invest 243:112-119.
Rigel DS (2010). Epidemiology of melanoma. Semin Cutan Med Surg 29:204-209.
Rochel N, Tocchini-Valentini G, Egea PF, Juntunen K, Garnier JM, Vihko P, et al. (2001). D. Functional and structural characterization of the insertion region in the ligand binding domain of the vitamin D nuclear receptor. Eur J Biochem 268:971 -979.
Ross AC, Manson JE, Abrams SA, Aloia JF, Brannon PM, Clinton SK, et al. (2011). The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: what clinicians need to know. J Clin Endocrinol Metab 96:53-58.
Schwartz GG, Whitlatch LW, Chen TC, Lokeshwar BL, Holick MF (1998). Human prostate cells synthesize 1,25-dihydroxyvitamin D3 from 25-hydroxyvitamin D3. Cancer Epidemiol Biomarkers Prev 7:391-395.
Shipman AR, Clark AB, Levell NJ (2011). Sunnier European countries have lower melanoma mortality. Clin Exp Dermatol 36:544-547.
Spina CS, Tangpricha V, Uskokovic M, Adorinic L, Maehr H, Holick MF (2006). Vitamin D and cancer. Anticancer Res 26:2515-2524.
Stambolsky P, Tabach Y, Fontemaggi G, Weisz L, Maor-Aloni R, Siegfried Z, et al. (2010). Modulation of the vitamin D3 response by cancer-associated mutant p53. Cancer Cell 17:273-285.
Stumpf WE, Sar M, Reid FA, Tanaka Y, De Luca HF (1 979). Target cells for 1,25-dihydroxyvitamin D3 in intestinal tract, stomach, kidney, skin, pituitary and parathyroid. Science 206:1188-1190.
Tagliabue E, Raimondi S, Gandini S (2015). Meta-analysis of vitamin D-binding protein and cancer risk. Cancer Epidemiol Biomarkers Prev 24:1 758-1 765.
Tang JY, Fu T, Leblanc E, Manson JE, Feldman D, Linos E, et al. (2011). Calcium plus vitamin D supplementation and the risk of nonmelanoma and melanoma skin cancer: post hoc analyses of the women’s health initiative randomized controlled trial. J Clin Oncol 29:3078-3084.
Uitterlinden AG, Fang Y, Van Meurs JB, Pols HA, Van Leeuwen JP (2004). Genetics and biology of vitamin D receptor polymorphisms. Gene 338:143-156.
Valverde P, Healy E, Jackson I, Rees JL, Thody AJ (1995). Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat Genet 11:328-330.
Vinceti M, Malagoli C, Fiorentini C, Longo C, Crespi CM, Albertini G, et al. (2011). Inverse association between dietary vitamin D and risk of cutaneous melanoma in a northern Italy population. Nutr Cancer 63:506-513.
Wang TJ, Zhang F, Richards JB, Kestenbaum B, van Meurs JB, Berry D, et al. (2010). Common genetic determinants of vitamin D insufficiency. Lancet 376:180-188.
Weinstein SJ, Purdue MP, Smith-Warner SA, Mondul AM, Black A, Ahn J, et al. (2015). Serum 25-hydroxyvitamin D, vitamin D binding protein and risk of colorectal cancer in the Prostate, Lung, Colorectal and Ovarian Cancer Screening Trial. Int J Cancer 136:654-664.
Welsh J (2011). Vitamin D metabolism in mammary gland and breast cancer. Mol Cell Endocrinol 347:55-60.
Wu XC, Eide MJ, King J, Saraiya M, Huang Y, Wiggins C, etal. (2011). Racial and ethnic variations in incidence and survival of cutaneous melanoma in the United States, 1999-2006. J Am Acad Dermatol 65:S26-S37.
Yuasa I, Umetsu K, Harihara S, Kido A, Miyoshi A, Saitou N, et al. (2007). Distribution of two Asian-related coding SNPs in the MC1R and OCA2 genes. Biochem Genet 45:535-542.
Zhao XZ, Yang BH, Yu GH, Liu SZ, Yuan ZY (2014). Polymorphisms in the vitamin D receptor (VDR) genes and skin cancer risk in European population: a metaanalysis. Arch Dermatol Res 306:545-553.
Zittermann A, Iodice S, Pilz S, Grant WB, Bagnardi V, Gandini S (2012). Vitamin D deficiency and mortality risk in the general population: a meta-analysis of prospective cohort studies. Am J Clin Nutr 95:91-100.