Vitamin D, Sunlight and Cancer Connection – Holick
Anti-Cancer Agents in Medicinal Chemistry Volume 13 Issue 1 pp.70-82 (13)
Author: Michael F. Holick
PDF with ALL graphics is attached at the bottom of this page
It has been more than 100 years when it was first appreciated that increased sun exposure reduced risk of dying of cancer. The most beneficial effect of sun exposure is the production of vitamin D in the skin. Recent evidence suggests that most cells in the body not only have a vitamin D receptor but also have the capacity to convert 25-hydroxyvitamin D to 1,25-dihydroxyvitamin D. Once formed 1,25-dihydroxyvitamin D can inhibit cellular proliferation, induce cellular maturation, inhibit angiogenesis and ultimately cause apoptosis to prevent malignancy. A multitude of studies have associated improved vitamin D status with decreased risk for developing several deadly cancers including colon, breast, pancreatic and ovarian cancers. Patients with cancer are at high risk for vitamin D deficiency. Sensible sun exposure, vitamin D fortification and vitamin D supplementation should be encouraged to improve the vitamin D status of children and adults not only for bone health but for reducing risk of developing and dying of cancer. The goal is to achieve a blood level of 25-hydroxyvitamin D of 40-60 ng/mL. This can be accomplished by children taking 600-1000 and adults 1500-2000 international units (IU) vitamin D daily from diet and supplements along with sensible sun exposure when the sun is capable of producing vitamin D in the skin.
Affiliation: Boston University School of Medicine, 85 East Newton Street, M-1013, Boston MA 02118.
INTRODUCTION
At the turn of the last century exposure to sunlight was heralded as a method of curing skin diseases including lupus vulgaris, a disease caused by a tuberculosis infection of the skin. This resulted in Finsen receiving the Nobel Prize in 1903 [1]. In 1921 Hess and Unger reported that exposure to sunlight was an effective treatment for rickets [2]. At approximately the same time Hoffman in 1915 compared cancer mortality in cities according to latitude between 1908 and 1912 and reported that cancer mortality increased in adults living farther from the equator [3]. Peller and Stephenson [4] observed a relationship between the morbidity and mortality of cancer and exposure to sunlight. They concluded that exposure to enough solar radiation caused a decrease in the incidence of malignant tumors in organs that were not at all accessible to treatment. They did however realize that this was accompanied by an increase in non-melanoma skin cancer which they considered as treatable. They analyzed the incidence of cancer in a population with increased exposure to solar UV radiation, i.e. U.S. Navy personnel aboard ships and observed that the rate of nonmelanoma skin cancer was 8 times higher in the Navy personnel while the total number of deaths from other cancers was 60% less than in the civilian population [4,5]. This was followed with the observation by Apperley [6] who analyzed the incidence and mortality of cancer in Northeastern and Southern states in the United States and in Canada and concluded that there was a highly significant correlation between reduced cancer mortality in adults who've engaged in agricultural activities outdoors. He also demonstrated that adults living in the Northeast were at higher risk for developing malignant deadly cancers compared to adults living in the South (Fig. 1).
In the 1980s-1990s the Garlands conducted several epidemiologic studies to evaluate the correlation between sun exposure, cancer and the vitamin D connection [7-9]. They demonstrated a strong negative correlation between latitude, increased sun exposure and vitamin D status and the risk of many different cancers including colon, breast, ovarian and melanoma [7-11]. These correlations were supported by the observation that breast and colon cancer mortality rates increased in cities with high levels of UV absorbing pollution thus preventing the vitamin D producing solar ultraviolet B (UVB) radiation from reaching the earth's surface [5]. These observations were followed by several other reports including those of Grant who observed a total of more than 13 cancers were reduced by adequate exposure to solar UVB radiation. He calculated in a span of 24 years, 1970-1994, a total of 566,400
Americans lost their lives due to cancer because of inadequate exposure to solar UVB radiation. He also estimated between 50,000-63,000 Americans and 19,000-25,000 British citizens in the United Kingdom die prematurely from cancer each year due to vitamin D deficiency [12-14].
VITAMIN D: SUNLIGHT AND DIETARY SOURCES
Humans have depended on sun exposure throughout evolution for their vitamin D requirement [15]. Vitamin D was essential for the development and maintenance of the vertebrate skeleton. During exposure to sunlight 7-dehydrocholesterol that is present in the plasma membrane of the living cells in the epidermis is converted to previtamin D3 [16]. Once formed previtamin D3 undergoes a thermally induced rearrangement of its double bonds to form vitamin D3. Vitamin D3 is ejected out of the plasma membrane into the extracellular space and diffuses into the dermal capillary bed for transport to the liver (Fig. 2).
The major natural dietary sources of vitamin D3 are from oily fish and cod liver oil (Table 1). Vitamin D2 that was discovered in the 1930s is obtained naturally from mushrooms exposed to sunlight and artificial ultraviolet radiation. Vitamin D2 is commercially produced from the ultraviolet irradiation of yeast while vitamin D3 is either obtained from fish oils or made from lanolin [17]. Various countries fortify some foods including dairy, cereals, orange juice and margarine with vitamin D. These countries include the United States, Canada, Mexico, Sweden and Finland. Typically there is no more than 100 international units (IU) in a serving. Most countries in Europe still forbid the fortification of foods with vitamin D. This is due to the misconception that an outbreak of hypercalcemia in young children in Great Britain was due to the over fortification of milk with vitamin D. However it is likely that these children suffered from a rare disease known as Williams syndrome. It is now recognized that the hypercalcemia observed in this disease is due to a hypersensitivity to vitamin D [18].
Fig. (1). Cancer Mortality per 100,000 population (white) and solar radiation index. (in 1941)
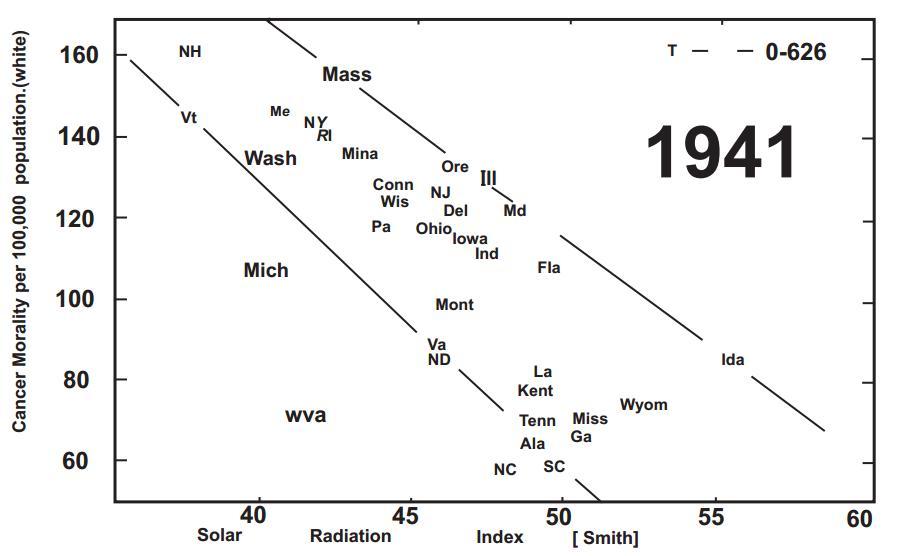
Copyright (1941), with permission from the American Association for Cancer Research.
VITAMIN D METABOLISM FOR CALCIUM HOMEOSTASIS AND BONE HEALTH
Once vitamin D (D represents either D2 or D3) is ingested from the diet or made in the skin it travels to the liver where it is metabolized to 25-hydroxyvitamin D [25(OH)D]. 25(OH)D is the major circulating form of vitamin D and is used by physicians to determine a patient's vitamin D status [17] (Fig. 2).
25(OH)D however is at physiologic concentrations biologically inactive on calcium and bone metabolism. It requires an additional hydroxylation on carbon-1 in the kidneys to form 1,25-dihydroxyvitamin D [1,25(OH)2D]. Once formed 1,25(OH)2D enters the circulation and travels into the small intestine where it interacts with the vitamin D receptor (VDR) unlocking genetic information that results in an increase in the efficiency of dietary calcium and phosphorus absorption [17]. It also travels to the skeleton and interacts with the VDR in the osteoblasts which provides a signal to monocytic osteoclasts to mature into multinucleated giant cells which are responsible for mobilizing calcium out of the skeleton to maintain calcium homeostasis [18 20].
VITAMIN D METABOLISM AND THE CANCER CONNECTION
By the mid-1990s it was generally accepted that living at higher latitude and being at higher risk for vitamin D deficiency was associated with increased risk of developing malignant tumors [514]. It was initially hypothesized that increased sun exposure or increased vitamin D intake resulted in an increase in the renal production of 1,25(OH)2D which in turn had anticancer properties. However this hypothesis was challenged because an increase in the production of 1,25(OH)2D would potentially cause hypercalcemia. Furthermore it was well-established that the renal production of 1,25(OH)2D was tightly regulated by calcium, phosphorus, parathyroid hormone and fibroblast growth factor 23 [17].
It had been known that activated macrophages [21] and cultured human keratinocytes [22] had the capacity to convert 25(OH)D3 to 1,25(OH)2D3. Schwartz and colleagues [23,24] in the mid-1990s reported that prostate cells grown from biopsies had the capacity to convert 25(OH)D3 to 1,25(OH)2D3 (Fig. 3). They further demonstrated that a prostate cancer cell line LNCaP which had a VDR lacked the 25-hydroxyvitamin D-1-hydroxylase (cyp 27B1;1-OHase) (Fig. 3). When these cells were incubated with 25(OH)D3 their proliferative activity was not affected and they were unable to produce 1,25(OH)2D3. However when the cells were transfected with the 1-OHase gene, 25(OH)D3 inhibited their (Fig. 4). It was also demonstrated that the cells converted 3H-25(OH)D3 to 3H-1,25(OH)2D3 [24]. At the same time several investigators reported that cells from the lung, breast, colon, brain among many other cell types also had the capacity to convert 25(OH)D3 to1,25(OH)2D3 [25-29]. Based on these observations it was concluded that the likely reason for why increased sun exposure and improved vitamin D status was associated with a reduced risk for deadly cancers was that higher circulating levels of 25(OH)D provided the substrate for the various tissues and organs that had a 1-OHase to locally produce 1,25(OH)2D. Once formed 1,25(OH)2D unlocks nuclear signals that could control cell growth and prevent malignancy, induce apoptosis and/or prevent angiogenesis [30-32]. 1,25(OH)D also enhances the expression of the 25-hydroxyvitamin D-24-hydroxylase (cyp24A1; 24-OHase). This enzyme causes the oxidation of the side chain to form a water-soluble biologically inactive calcitroic acid (Fig. 2).
ANTIPROLIFERATIVE ACTIVITY OF 1,25(OH)D3 AND NALOGS
It is estimated that more than 2000 genes may be directly or indirectly regulated by 1,25(OH)2D [20,21,33]. In the early 1980s it was first appreciated that leukemic cells that had a VDR. When they were exposed to 1,25(OH)2D3 their proliferation was inhibited and the cells matured into normal appearing macrophages [34,35]. This was followed by the observation that 1,25(OH)2D3 inhibited human keratinocyte proliferation and induced terminal differentiation [36]. These exciting observations were immediately tested for their clinical utility in treating leukemia [37] and the
Fig. (2).
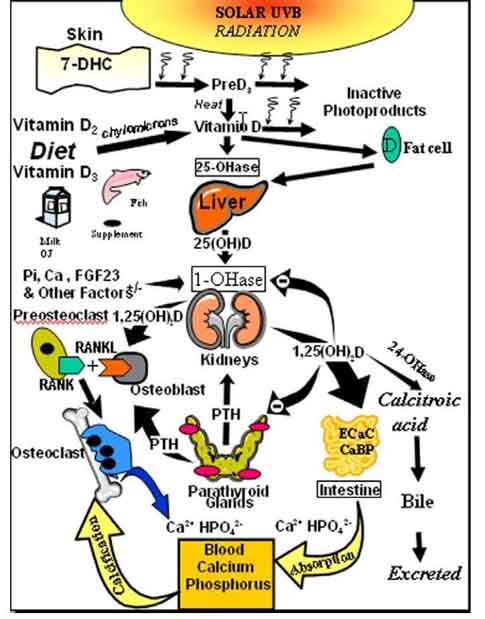
Schematic representation of the synthesis and metabolism of vitamin D for regulating calcium, phosphorus and bone metabolism. During exposure to sunlight 7-dehydrocholesterol in the skin is converted to previtamin D3. PreD3 immediately converts by a heat dependent process to vitamin D3. Excessive exposure to sunlight degrades previtamin D3 and vitamin D3 into inactive photoproducts. Vitamin D2 and vitamin D3 from dietary sources is incorporated into chylomicrons, transported by the lymphatic system into the venus circulation. Vitamin D (D represents D2 or D3) made in the skin or ingested in the diet can be stored in and then released from fat cells. Vitamin D in the circulation is bound to the vitamin D binding protein which transports it to the liver where vitamin D is converted by the vitamin D-25-hydroxylase to 25-hydroxyvitamin D [25(OH)D]. This is the major circulating form of vitamin D that is used by clinicians to measure vitamin D status (although most reference laboratories report the normal range to be 20-100 ng/ml, the preferred healthful range is 30-60 ng/ml). It is biologically inactive and must be converted in the kidneys by the 25-hydroxyvitamin D-1a-hydroxylase (1-OHase) to its biologically active form 1,25-dihydroxyvitamin D [1,25(OH)2D]. Serum phosphorus, calcium fibroblast growth factors (FGF-23) and other factors can either increase (+) or decrease (-) the renal production of 1,25(OH)2D. 1,25(OH)2D feedback regulates its own synthesis and decreases the synthesis and secretion of parathyroid hormone (PTH) in the parathyroid glands. 1,25(OH)2D increases the expression of the 25-hydroxyvitamin D-24-hydroxylase (24-OHase) to catabolize 1,25(OH)2D to the water soluble biologically inactive calcitroic acid which is excreted in the bile. 1,25(OH)2D enhances intestinal calcium absorption in the small intestine by stimulating the expression of the epithelial calcium channel (ECaC) and the calbindin 9K (calcium binding protein; CaBP). 1,25(OH)2D is recognized by its receptor in osteoblasts causing an increase in the expression of receptor activator of NFkB ligand (RANKL). Its receptor RANK on the preosteoclast binds RANKL which induces the preosteoclast to become a mature osteoclast. The mature osteoclast removes calcium and phosphorus from the bone to maintain blood calcium and phosphorus levels. Adequate calcium and phosphorus levels promote the mineralization of the skeleton. Holick copyright 2007. Reproduced with permission.
hyperproliferative skin disorder psoriasis [38]. Unfortunately patients with leukemia developed hypercalcemia and went into blastic phase and died [37]. Psoriasis patients treated with topical [38] or oral 1,25(OH)2D3 [39] responded well to therapy without developing hypercalcemia. Since these initial clinical studies several other studies had been conducted with 1,25(OH)2D3 and its analogs to treat liver carcinoma and prostate cancer [40-42]. These studies were halted due either to hypercalcemia or lack of efficacy. 1,25(OH)2D3 and its analogs have been effective in treating psoriasis and remain a first line treatment for this enigmatic hyperproliferative skin disorder [38,39,42-44].
Table 1. Sources of Vitamin D2 and Vitamin D3 (with Permission, Copyright Holick 2007)
MECHANISMS FOR 1,25(OH)D3 TO INHIBIT TUMOR GROWTH
Although the exact mechanisms by which 1,25(OH)2D3 regulates cellular proliferation and differentiation is not fully understood it is known that 1,25(OH)2D3 can control cell replication by regulating cell cycle genes including p27, p21 and p53 [30,40,45-47]. The proteins bind and inactivate cyclin dependent kinase complexes in the G1 phase causing the cell to remain in the G1 phase of the cell cycle. 1,25(OH)2D3 also decreases p27 degradation and its downstream effect is to inhibit cellular proliferation, cell migration and apoptosis. Similarly 1,25(OH)2D3 increases the expression of p21 which can induce apoptosis and G2/M cell cycle arrest [45-47] (Fig. 5) 1,25(OH)2D3 has been demonstrated to inhibit proliferation of endothelial cells and decrease VEGF expression. The VDR knockout mice have increased blood vessel size and volume compared to wild type mice [48]. Therefore it has been suggested that vitamin D through its local conversion of 25(OH)D to 1,25(OH)2D is responsible for controlling genes that keep cells in a normal proliferative state.
Fig. (3).
The 1 a-OHase activities in primary cultures of normal, BPH, and prostate cancer cells (CaP), and in human prostate cancer cell lines, DU145, PC-3 and LNCaP cells. Bars shown are standard deviation of three determinations. The four prostate cancer cultures were obtained from P1 (63y Caucasian), P2 (50y African-American), P3 (67y Caucasian), and P4 (53y Caucasian) prostate cancer patients. Three normal cultures were obtained from histologically normal prostates of a 21 and a 27-year-old donors and a 42-year-old African-American organ donor. BPH cultures were derived from an open prostatectomy specimen of a 58-year-old Caucasian and from a 60-year-old Caucasian. Reproduced with permission (24).
Fig. (4).
Effect of 25-hydroxyvitamin D3 [25(OH)D3] (10-8M) on the incorporation of 3H-thymidine into DNA of LNCaP cells transfected with cDNA encoding 25(OH)D-1 a-hydroxylase (1 a-OHase). (a) LNCaP cells were transfected transiently with PCR 3.1 vector, antisense (AS) or sense (S) 1 a-OHase cDNA. (b) LNCaP cells were stably transfected with either PCR 3.1 vector or with sense 1a-OHase cDNA. Data are presented as % of mock transfected control in the absence of 25(OH)D3. Data are mean + SD, n=8, *P<0.05. Reproduced with permission (24). However if the cell becomes hyperproliferative 1,25(OH)2D can induce cell death or alternatively inhibit angiogenesis to prevent a nutrient supply to the malignant cell preventing its proliferation.
VITAMIN D NUTRITION AND CANCER IN WOMEN, MEN AND MICE
There is overwhelming scientific evidence suggesting that vitamin D sufficiency is important for the prevention of a wide variety of deadly cancers. Woo et al. [49] reported that men with metastatic prostate cancer had as much as a 50% decrease in their PSA in response to 2000 IU/d vitamin D3 for up to 21 months. Lappe et. al. [50] reported a more than 60% reduction in the development of all cancers in a small study of postmenopausal women who received calcium supplementation along with 1100 IU/d (Fig. 6) [50]. The Women's Health Initiative (WHI) reported that calcium and vitamin D supplementation in postmenopausal women did not influence the incidence of colorectal cancer. However women who had a blood level of 25(OH)D <12 ng/mL (to convert
Fig. (5).
Vitamin D maintains cellular growth by controlling several genes that control cellular proliferation and differentiation. 25-hydroxyvitamin D [25 (OH)D] is converted to 1,25-dihydroxyvitamin D [1,25(OH)2D] in a wide variety of non-renal cells including cells in the colon and prostate. 1,25(OH)2D interacts with the vitamin D receptor (VDR) and regulates a variety of genes that control apoptosis, proliferation and differentiation. Reproduced with permission; Holick copyright 2009.
ng/mL into nmol/L multiply by 2.496 ) had a 253% increased risk for developing colorectal cancer during the 8 years of study compared to women who were vitamin D sufficient at the beginning of the study and had a blood level of 25(OH)D of 24 ng/mL [51]. Further a subgroup analysis restricted to women not on personal calcium and/or vitamin D suggested that the women who took the calcium 1000 mg and 400 IU vitamin D3 daily for 8 years had a 14-20% reduced risk for developing breast cancer and a 17% reduced risk for developing colorectal cancer [52]. The importance of vitamin D in reducing risk of colorectal cancer is also supported by the observation that the vitamin D receptor polymorphisms was associated with colorectal cancer and in the same study a 34% reduced risk of developing colorectal cancer was seen in adults who had blood levels of 25(OH)D of 31-34 ng/mL [53]. A recent meta-analysis for the US Preventative Services Task Force regarding vitamin D supplementation concluded that for each 4 ng/mL increase in blood 25(OH)D concentrations was associated with a 6% (95% Cl 3-9%) reduced risk for colorectal cancer [54].
Animal studies have also supported the concept that an adequate amount of nutritional vitamin D reduces tumor growth. Tangpricha et al. [55] observed that mice who received an adequate amount of vitamin D in their diet to maintain their blood levels of 25(OH)D of 26 ±6 ng/mL had a 40% reduction in the tumor size of a mouse colon cancer compared to mice who were on a vitamin D deficient diet and had undetectable levels of 25(OH)D. (Fig. 7) To determine if vitamin D nutrition could also influence a human tumor's growth, a study was conducted in athymic mice that were injected subcutaneously with an androgen insensitive prostate cancer cell line DU-145. The tumor volume was dramatically reduced by more than 50% in the mice that had adequate vitamin D in their diet and had maintained a blood level of 25(OH)D of 28.1±2.2 ng/mL compared to the mice on a normal calcium vitamin D deficient diet that had a blood level of 25(OH)D that was undetectable [56].
Fig. (6).
Kaplan-Meier survival curves (ie, free of cancer) for the 3 treatment groups ramdomly assigned in the cohort of women who were free of cancer at 1 y of intervention (n = 1085). Sample sizes are 266 for the placebo group, 416 for the calcium-only (Ca-only) group, and 403 for the calcium plus vitamin D (Ca+D) group. The survival at the end of study for the Ca + D group is significantly higher than that for the placebo group, by logistic regression. Reproduced with permission from Dr. Robert Heaney 2007.
CANCER RESISTANCE TO VITAMIN D
Several studies have reported that cancer patients are at high risk for being vitamin D deficient [30,41,57,58]. This is due in part to them being advised to avoid sun exposure because of their chemotherapy. In addition they often have gastrointestinal side effects including nausea from the chemotherapy reducing dietary vitamin D intake and causing vitamin D malabsorption. Therefore it
Fig. (7).
Effect of Vitamin D Status on MC-26 tumor size in vivo. Six week old Balb/c mice were randomized into a vitamin D deficient normal calcium (0.47%) diet (n= 9) (solid line, black diamonds) and vitamin D sufficient (n=10) (500,000 IU/kg diet) and normal calcium (0.47%) diet (dashed line, black triangles). When the vitamin D deficient mice had serum 25(OH)D levels of < 12.5 nmol/L, mice in each group was injected with 10,000 cells of MC-26. Mice were sacrificed at day 20. Results are mean tumor volume ± SE in each group. Repeated measures ANOVA was significant at p=0.002. Post-hoc analysis with two sample student's t-tests demonstrated significant differences between groups starting at day 9 (noted by *). Reproduced with permission (55).
is important to correct the vitamin D nutritional deficiency in patients who have cancer. The vitamin D may help improve mood, nonspecific muscle aches and pains and could potentially enhance the therapeutic efficacy of the chemotherapy along with improving overall feeling of wellbeing.
However vitamin D by itself is not a treatment for cancer. The role of vitamin D in reducing risk and growth of cancers is due to its pleotrophic effects on regulating cellular proliferation, differentiation, apoptosis and angiogenesis once 25(OH)D is converted to 1,25(OH)2D [59,60]. Unfortunately cancer cells have found a variety of mechanisms to reduce vitamin D's effectiveness in fighting the malignancy. First and foremost the cancer can increase the destruction of 1,25(OH)2D by inducing the cyp24a1 which results in the oxidation of the side chain into the water-soluble calcitroic acid. Human prostate cancer cells that have robust expression of the cyp24a1 are unresponsive to the anti-proliferative activity of 1,25(OH)2D [40,47]. Malignant colon cancer cells also express the transcription factor SNAIL which is important for cell movement and exists in both invertebrates and vertebrates. SNAIL also binds VDR and prevents it from acting as a transcription factor to regulate cellular proliferation and differentiation. By doing so it prevents 1,25(OH)2D- VDR complex from binding to beta-catenin which is a known factor that enhances cellular proliferation. SNAIL1 was also shown to also inhibit expression of VDR and E-cadherin which is important for cellular adhesion and differentiation [61] (Fig. 8).
CONCLUSION
There are several thousand publications that have associated sunlight and vitamin D deficiency with increased risk for developing and dying of a wide variety of cancers [62-65]. The fact that the VDR exists in most cells in the body and that the cells have the capability of producing 1,25(OH)2D provides a strong rationale to support the hypothesis that maintaining a healthy vitamin D status with a 25(OH)D in the desired range of 40-60 ng/mL,as recommended by the Endocrine Society's Practice Guidelines [66], may reduce risk for malignancy and improve survival rates for several cancers [58,67-70].
The major source of vitamin D for most children and adults is exposure to sunlight. Unfortunately for more than 40 years has been the message to avoid all direct sun exposure because of concern for developing "skin cancer" [71,72]. There is no debate that excessive sun exposure especially on the face and dorsum of the hands increases risk for nonmelanoma skin cancers including basal cell and squamous cell carcinomas [73]. These cancers are easy to detect and easy to treat and if detected early and appropriately treated are curable. However what is of great concern is the deadly skin cancer melanoma. Unlike the non-melanoma skin cancers most melanomas occur on the least sun exposed areas [73]. More importantly occupational sun exposure has been associated with a reduced risk for melanoma [73,74]. In the 'skin cancer capital' Australia, it has been estimated that more than 40% of the population is now at risk for vitamin D deficiency [75]. A survey of Australian dermatologists in the summer revealed that 87% had a blood level of 25(OH)D <20 ng/mL [76]. The sun-safe message in Australia and New Zealand had been modified in part due to recognizing the widespread consequence which is vitamin D deficiency and insufficiency. The New Zealand Bone and Mineral Society in cooperation with the Australian College of Dermatologists and the Cancer Council for Australia in 2008 had recommended a balance between avoiding the increased risk for skin cancer and achieving enough ultraviolet radiation to maintain adequate vitamin D levels [30]. This message was reinforced by the 2012 Consensus Statement on Vitamin D and Sun exposure in New Zealand that Advice on sun exposure requires balancing the risk of skin damage and skin cancer against the risk of vitamin D deficiency.
They recommended
For the general population, some sun exposure is recommended for vitamin D synthesis.
For older adults who are mobile and living independently, the same sun safety messages apply as for the general population. Physical activity outdoors should be encouraged.
•*Physical activity is associated with increased vitamin D levels.
Being active while outside may enable more skin to be exposed, increase vitamin D production and reduce the length of time required for vitamin D synthesis." [77] The Canadian Cancer Society which now recommends that all Canadian adults ingest 2000 IU/d of vitamin D. In addition the Canadian Dermatologic Association, National Council on Skin Cancer Prevention (US) and the World Health Organization Collaborative Center for the Promotion of Sun Protection are recognizing the need to minimize the health risks associated with excessive ultraviolet B radiation exposure while maximizing the potential benefits of optimum vitamin D status, supplementation, and small amounts of sun exposure for obtaining adequate vitamin D. It has been estimated that an adult in a bathing suit receiving an amount of sun exposure that causes a slight pinkness to the skin 24 hours later (one minimal erythemal dose; MED) is equivalent to
Fig. (8).
Top, micrographs of SNAIL-HA and mock-infected cells. Arrows indicate the phenotypic change induced by SNAIL. Bar, 50 urn. Below, immunostaining of ectopic SNAIL expression using an antibody to HA. Bar, 10 urn. Left normalized SNAIL. VDR and E-cadherin mRNA levels were measured by real-time RT-PCR. Right, protein expression was estimated by western blot. Numbers refer to fold increase over untreated mock-infected cells. SNAIL inhibits the induction of L1-NCAM and filamin by 1,25(OH)D2D3. Wild-type (left) but not mutant (right) SNAIL proteins inhibit VDR transcriptional activity (4XVDRE-tk-luciferase). Reproduced with permission (59).
Fig. (9).
Comparison of serum vitamin D3 levels after a whole-body exposure (in a bathing suit; bikini for women) to 1 MED (minimal erythemal dose) of simulated sunlight compared with a single oral dose of either 10,000 or 25,000 IU of vitamin D2. Holick copyright 2004. Reproduced with permission.
Fig. (10).
Adults age 18 - 65 exposed to UVB radiation three times a week for three months or receiving 1000 IU of vitamin D2 or vitamin D3 daily. Holick copyright 2008. Reproduced with permission.
ingesting a single dose of 20,000 IU of vitamin D (Fig. 9). Exposure of healthy adults with skin types 2, 3 and 4 to 0.5 MED once a week for 3 months was more effective in maintaining serum 25(OH)D levels than taking an oral dose of 1000 IU vitamin D3 daily (Fig. 10). Furthermore it has been demonstrated that vitamin D produced in the skin from sun exposure last 2-3 times longer in the circulation compared to when it is taken as an oral supplement [78].
Although a few studies have suggested that raising blood levels of 25(OH)D above 30 ng/mL increases risk for all-cause mortality [79] and prostate cancer [80] a preponderance of studies have not supported these observations. There has been a lot of discussion about the so-called J curve regarding mortality and vitamin D status. It was suggested that elderly adults with blood levels of 25(OH)D >30 ng/mL were at higher risk for mortality [79]. However one of the studies used in the analysis suggested that mortality continued to be reduced in both men and women until a blood level of 25(OH)D reached 50 ng/mL and that there may have been a slight increase in mortality above this level and only in women not men [81]. Thomas et al. [82] recently reported improvement in vitamin D status reduced risk for all-cause and cardiovascular mortality. In this prospective study 1801 adults with documented metabolic syndrome and after adjusting for confounders they observed a 75% reduction in all-cause mortality and 66% reduction in cardiovascular disease mortality with 85% and 76% reductions in mortality associated with congestive heart
Fig. (11).
A Schematic Representation of the Major Causes for Vitamin D Deficiency and Potential Health Consequences. Holick copyright 2010. Reproduced with permission.
Table 2. Vitamin D Intakes Recommended by the IOM and the Endocrine Practice Guidelines Committee
failure and sudden death. These data support recent meta-analyses showing that low 25(OH)D levels are a risk factor for mortality in CKD patients and in general populations. The lowest mortality risk in general populations was observed at 25(OH)D levels between 30 to 35 ng/mL [83,84]. A meta-analysis of RCTs confirmed that vitamin D supplementation significantly reduces all-cause mortality [85].
Not only will enhancing a child's and adult's vitamin D status reduce risk for developing cancer including lymphoma, prostate and breast cancer among many others [86-90] but will also reduce risk of other chronic illnesses including autoimmune diseases such as rheumatoid arthritis, multiple sclerosis and type 1 diabetes, infectious diseases, type 2 diabetes and cardiovascular disease (Fig. 11). Unless the person has a hypersensitivity to vitamin D due to a granulomatous disorder such as sarcoidosis or has a lymphoma that is capable of producing 1,25(OH)2D in an unregulated fashion there is no downside to increasing a person's vitamin D intake [66]. Children during their first year can ingest 400-1000 IU/d, children one year and older require 600-1000 IU/d and all adults should obtain from their diet and supplements 1500-2000 IU/d. Obese adults with a BMI >30 and children and adults on glucocorticoids, AIDS medications and anti-seizure medications may require 2-3 times more vitamin D to satisfy their requirement and to maintain blood levels 40-60 ng/ml (Table 2). Blood levels up to 100 ng/mL is considered to be safe [66,91].
SOURCES OF SUPPORT
Supported in part by the UV Foundation and the Mushroom Council.
CONFLICTS OF INTEREST
The author confirms that this article content has no conflicts of interest.
ACKNOWLEDGEMENTS
Declared none.
REFERENCES
[1] Holick, M.F. In: Biologic effects of light: Historical and new perspectives, Proceedings of a symposium, Basel, Switzerland November 1-3, 1998; Holick MF, Jung EG, Eds.; Kluwer Academic Publishers: Boston, MA, 1999. pp 11-32.
[2] Hess, A.F.; Unger, L.J. The cure of infantile rickets by sunlight. J. Am. Med. Assoc., 1921, 77, 39-41.
[3] Hoffman, F.L. The mortality of cancer throughout the world. Appendix E, Prudential Press. 1915.
[4] Peller, S.; Stephenson, C.S. Skin irritation and cancer in the United States Navy. Am. J. M. Sc., 1937. 194, 326-333.
[5] Spina, C., Tangpricha, V., Yao, M., Zhou, W., Wolfe, M.M., Maehr, H., Uskokovic, M., Adorini, L., and Holick, M.F. Colon cancer and solar ultraviolet B radiation and prevention and treatment of colon cancer in mice with vitamin D and its Gemini analogs. J. Steroid Biochem & Molecular Biol., 2005. 97, 111-120.
[6] Apperly, F.L. The relation of solar radiation to cancer mortality in North America. Cancer Research. 1941. 1, 191-195.
[7] Garland, C.; Shekelle, R.B.; Barrett-Connor, E.; Criqui, M.H.; Rossof, A.H.; Oglesby, P. Dietary vitamin D and calcium and risk of colorectal cancer: A 19-year prospective study in men. Lancet. 1985. 9, 307-309.
[8] Garland, C.F.; Garland, F.C.; Shaw, E.K.; Comstock, G.W.; Helsing, K.J.; Gorham, E.D. Serum 25-hydroxyvitamin D and colon cancer: Eight-year prospective study. Lancet. 1989.18, 11761178.
[9] Garland, C.F.; Garland, F.C.; Gorham, E.D.; et al. The role of vitamin D in cancer prevention. Am. J. Public Health, 2006. 96(2), 252-61.
[10] Garland, F.C.; Garland, C.F. Occupational sunlight exposure and melanoma in the U.S. Navy. Arch. Env. Health, 1990. 45, 261-267.
[11] Garland, C.F.; Gorham, E.D.; Mohr, S.B.; Grant, W.B.; Giovannucci, E.L.; Lipkin, M.; Newmark, H.; Holick, M.F.; Garland, F.C. Vitamin D and prevention of breast cancer: Pooled analysis. J. SteroidBiochem. Mol. Biol., 2007. 103(3-5), 708-11.
[12] Grant, W.B. An estimate of premature cancer mortality in the U.S. due to inadequate doses of solar ultraviolet-B radiation. Cancer. 2002. 70, 2861-2869.
[13] Grant, W.B. An estimate of the global reduction in mortality rates through doubling vitamin D levels. Eur. J. Clin. Nutr., 2011. 65(9), 1016-26.
[14] Grant, W.B. Lower vitamin-D production from solar ultraviolet-B Irradiance may explain some differences in cancer survival rates. J. National Medical Association. 2006. 98(3), 357-64.
[15] Holick, M.F. Health Benefits of Vitamin D and Sunlight: A D-bate. Nat. Rev. Endocrinol., 2011. 7, 73-75.
[16] Holick, M.F.; MacLaughlin, J.A.; Clark, M.B.; Holick, S.A.; Potts, J.T. Jr; Anderson, R.R.; Blank, I.H.; Parrish, J.A.; Elias, P. Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science, 1980. 210, 203-205.
[17] Holick, M.F. Vitamin D Deficiency. N. Eng. J. Med., 2007. 357, 266-281.
[18] Holick, M.F. The D-Lightful Vitamin D for Child Health. J. Parenter Enteral. Nutr., 2012. 36, 9S-19S.
[19] Khosla, S. The OPG/RANKL/RANK system. Endocrinol., 2001. 142(12), 5050-5055.
[20] Holick, M.F. Vitamin D and Health: Evolution, Biologic Functions, and Recommended Dietary Intakes for Vitamin D. Clin. Rev. Bone Miner. Metab, 2009. 7(1), 2-19.
[21] Adams, J.S.; Hewison, M. Update in Vitamin D. J. Clin. Endocrinol. Metab, 2010. 95(2), 471-478.
[22] Bikle, D. D.; Nemanic, M.D.; Whitney, J.O.; Elias, P.O. Neonatal human foreskin keratinocytes produce 1,25-dihydroxyvitamin D3. Biochemistry, 1986. 25, 1545-1548.
[23] Schwartz, G.G.; Whitlatch, L.W.; Chen, T.C.; Lokeshwar, B.L.; Holick, M.F. Human prostate cells synthesize 1,25-dihydroxyvitamin D3 from 25-hydroxyvitamin D3. Cancer Epidemiol Biomarkers Prev., 1998. 7, 391-395.
[24] Whitlatch, L.W.; Young, M.V.; Schwartz, G.G.; Flanagan, J.N.; Burnstein, K.L.; Lokeshwar, B.L.; Rich, E.S.; Holick, M.F.; Chen,T.C. 25-hydroxyvitamin D-1-a-hydroxylase activity is diminished in human prostate cancer cells and is enhanced by gene transfer. J. Steroidiochem. Mol. Biol., 2002. 81(2), 135-140.
[25] Krishnan, A.V.; Peehl, D.M.; Feldman, D. The role of vitamin D in prostate cancer. Recent Results Cancer Res., 2003. 164, 205-221.
[26] Mawer, E.B.; Hayes, M.E.; Heys, S.E.; Davies, M.; White, A.; Stewart, M.F.; Smith, G.N. Constitutive synthesis of 1,25-dihhdroxyvitamin D3 by a human small cell lung cell line. J. Clin. Endocrinol. Metab., 1994. 79(2), 554-560.
[27] Radermacher, J.; Diesel, B.; Seifert, M.; Tilgen, W.; Reichrath, J.; Fischer, U.; Meese, E. Expression Analysis of CYP27B1 in Tumor Biopsies and Cell Cultures. Anticancer Research, 2006. 26, 2683-2686.
[28] Tangpricha, V.; Flanagan, J.N.; Whitlatch, L.W.; Tseng, C.C.; Chen, T.C.; Holt, R.P.; Lipkin, M.S.; Holick, M.F. 25-hydroxyvitamin D-1 a-hydroxylase in normal and malignant colon tissue. Lancet, 2001. 357(9269), 1673-1674.
[29] Cross, H.S.; Kallay, E.; Lechner, D.; Gerdenitsch, W.; Aldercreutz, H.; Armbrecht, H.J. Phytoestrogens and vitamin D metabolism: a new concept for the prevention and therapy of colorectal, prostate and mammary carcinoma. J. Nutr., 2004. 134(5), 1207S-1212S.
[30] Holick, M.F. Vitamin D and sunlight: strategies for cancer prevention and other health benefits. Clin. J. Am. Soc. Nephrol., 2008, 3(5), 1548-54.
[31] Guzey, M.; Kitada, S.; Reed, J.C. Apoptosis Induction by 1a,25-Dihydroxyvitamin D3 in Prostate Cancer. Mol. Can. Therapeut, 2002. 1, 667-677.
[32] Mantell, D.J.; Owens, P.E.; Bundred, N.J.; Mawer, E.B.; Canfield, A.E. 1 a,25-dihydroxyvitamin D3 inhibits angiogenesis in vitro and in vivo. Circ. Res., 2000. 87, 412-220.
[33] Nagpal, S.; Na, S.; Rathnachalam, R. Noncalcemic actions of vitamin D receptor ligands. Endocr. Rev., 2005. 26, 662-87.
[34] Abe, E.; Miyaura, C.; Sakagami, H.; Suda, T. Differentiation of rat myc leukemic cells by 1,25-dihydroxyvitamin D3. Proc. Natl. Acad. Sci. USA, 1981. 78, 4990-4994.
[35] Honma, Y.; Hozumi, M.; Abe, E; Konno, K.; Fukushima, M.; Hata, S.; Nishii, Y.; DeLuca, H. F. 1y,25-Dihydroxyvitamin D3 and 1y-hydroxyvitamin D3 prolong survival time of mice inoculated with myeloid leukemia cells. Proc. Natl. Acad Sci. USA, 1982. 80, 201-204.
[36] Smith, E.L.; Walworth, N.D.; Holick, M.F. Effect of 1,25-dihydroxyvitamin D3 on the morphologic and biochemical differentiation of cultured human epidermal keratinocytes grown in serum-free conditions. J. Invest. Dermatol., 1986. 86, 709-714.
[37] Koeffler, H.P.; Hirjik, J.; Iti, L.; the Southern California Leukemia Group. 1,25-Dihydroxyvitamin D3: in vivo and in vitro effects on human preleukemic and leukemic cells. Cancer Treat. Rep., 1985. 69, 1399-1407.
[38] Perez, A.; Chen, T.C.; Turner, A.; Raab, R.; Bhawan, J.; Poche, P.; Holick, M.F. Efficacy and safety of topical calcitriol (1,25-dihydroxyvitamin D3) for the treatment of psoriasis. Brit. J. Dermatol., 1996, 134, 238-246.
[39] Perez, A.; Raab, R.; Chen, T. C.; Turner, A.; Holick, M.F. Safety and efficacy of oral calcitriol (1,25-dihydroxyvitamin D3) for the treatment of psoriasis. Brit J. Dermatol., 1996, 134, 1070-1078.
[40] Feldman, D.; Zhao, X.Y.; Krishnan, A.V. Editorial/Mini-review: Vitamin D and prostate cancer. Endocrinology, 2000, 141 , 5-9.
[41] Beer, T.M.; Ryan, C.W.; Venner, P.M.; Petrylak, D.P.; Chatta, G.S.; Ruether, J.D.; Redfern, C.H.; Fehrenbacher, L.; Saleh, M.N.; Waterhouse, D.M.; Carducci, M.A.; Vacario, D.; Dreicer, R.; Higano, C.S.; Ahmann, F.R.; Chi, K.N.; Henner, W.D.; Arroyo, A.; Clow, F.W. Double-blinded randomized study of igh-dose calcitriol plus docetaxel compared with placebo plus docetaxel in androgen-independent prostate cancer: a report from the ASCENT investigators. J. Clin. Oncol., 2007, 25, 669-674.
[42] Dalhoff, K.; Dancey, J.; Astrup, L.; et al. A phase II study of the vitamin D analogue Seocalcitol in patients with inoperable hepatocellular carcinoma. Br. J. Cancer, 2003, 89, 252-57.
[43] Kragballe, K. Treatment of psoriasis by the topical application of the novel vitamin D3 analogue MC 903. Arch. Dermatol, 1989. 125, 1647-1652.
[44] Durakovic, C.; Malabanan, A.; Holick, M. F. Rationale for use and clinical responsiveness of hexafluoro-1,25-dihydroxyvitamin D3 for the treatment of plaque psoriasis: A pilot study. Brit J. Dermatol. , 2001, 144(3), 500-506.
[45] Saramaki, A.; Banwell, C.M.; Canpbell, M.J.; Carlberg, C. Regulation of the human p21(waf1/cip1) gene promoter via multiple binding sites for p53 and the vitamin D3 receptor. Nucleic Acids Res., 2006, 34, 543-554.
[46] Lambert, J.R.; Kelly, J.A.; Shim, M.; Huffer, W.E.; Nordeen, S.K.; Baek, S.J.; Eling, T.E.; Lucia, M.S. Prostate Derived Factor in Human Prostate Cancer Cells: Gene Induction by Vitamin D via a p53-Dependent Mechanism and Inhibition of Prostate Cancer Cell Growth. J. Cell Physiol., 2006, 208, 566-574.
[47] Chen, T.C.; Holick, M.F. Vitamin D and prostate cancer prevention and treatment. Trends in Endocrinol Metabol., 2003, 14, 423-430.
[48] Chung, I.; Han, G.; Seshadri, M.; et al. Role of Vitamin D Receptor in the Antiproliferative Effects of Calcitriol in Tumo-Derived Endothelial Cells and Tumor Angiogenesis In vivo. Cancer Res., 2009, 69, 967-975.
[49] Woo, T.C.S.; Choo, R.; Jamieson, M.; Chander, S.; Vieth R. Pilot Study: potential role of vitamin D (cholecalciferol) in patients with PSA relapse after definitve therapy. Nutrition and Cancer, 2005, 51(1), 32-36.
[50] Lappe, J.M.; Travers-Gustafson, D.; Davies, K.M.; Recker, R.R.; Heaney, R.P. Vitamin D and calcium supplementation reduces cancer risk: results of a randomized trial. Am. J. Clin. Nutr., 2007, 85(6), 1586-1591.
[51] Holick, M.F. Calcium plus Vitamin D and the Risk of Colorectal Cancer. N. Engl. J. Med., 2006, 354(21), 2287-2288.
[52] Bolland, M.J.; Grey, A.; Gamble, G.D.; Reid, I.R. Calcium and vitamin D supplements and health outcomes: a reanalysis of the Women's Health Initiative (WHI) limited-access data set. Am. J. Clin. Nutr., 2011, 94, 1144-9.
[53] Yamaji, T.; Iwasaki, M.; Sasazuki, S.; Sakamoto, H.; Yoshida, T.; Tsugane, S. Association Between Plasma 25-Hydroxyvitamin D and Colorectal Adenoma According to Dietary Calcium Intake and Vitamin D Receptor Polymorphism. Am. J. Epidemiol., 2012, 175(3), 236-244.
[54] Chung, M.; Lee, J.; Terasawa, T.; Lau, J.; Trikalinos, T.A. Vitamin D With or Without Calcium Supplementation for Prevention of Cancer and Fractures: An Updated Meta-analysis for the U.S. Preventive Services Task Force. Ann. Intern. Med., 2011, 155, 827838.
[55] Tangpricha, V.; Spina, C.; Yao, M.; Chen, T.C.; Wolfe, M.M.; Holick, M.F. Vitamin D deficiency enhances the growth of MC-26 colon cancer xenografts in Balb/c mice. J. Nutr., 2005, 135(10), 2350-2354.
[56] Ray, R.; Banks, M.; Abuzahra, H.; Eddy, V.J.; Persons, K.S.; Lucia, M.S.; Lambert, J.R.; Holick, M.F. Effect of Dietary Vitamin D and Calcium on the Growth of Androgen-insenstive Human Prostate Tumor in a Murine Model. Anticancer Res., 2012, 32, 727-732.
[57] Tangpricha, V.; Colon, N.A.; Kaul, H.; Wang, S.L.; Decastro S.; Blanchard, R.; Chen, T.; Holick, M.F. Prevalence of vitamin D deficiency in patients attending an outpatient cancer care clinic inBoston. Endo. Prac., 2004, 10(3), 292.
[58] Ng, K.; Meyerhardt, J.A.; Wu, K.; Feskanich, D.; Hollis, B.W.; et al. Circulating 25-hydroxyvitamin D levels and survival in patients with colorectal cancer. J. Clin. Oncol., 2008, 26, 2984-91
[59] Fleet, J.C.; Desmet, M.; Johnson, R.; Li, Y.; Vitamin D and cancer: a review of molecular mechanisms. Biochem. J., 2012, 441(1), 61-76.
[60] Krishnan, A.V.; Feldman, D. Mechanisms of the anti-cancer and anti-inflammatory actions of vitamin D. Annu. Rev. Pharmacol. Toxicol., 2011, 51, 311-36.
[61] Palmer, H.G.; Larriba, M.J.; Garcia, J.M.; Ordonez-Moran, P.; Pena, C.; Peiro, S.; Puig, I.; Rodriguez, R.; de la Fuente, R.; Bernad, A.; Pollan, M.; Bonilla, F.; Camallo, C.; Garcia de Herreros, A.; Munoz, A. The transcription factor SNAIL represses vitamin D receptor expression and responsiveness in human colon cancer. Nature Medicine, 2004, 10, 917-919.
[62]PROBLEMS WITH EXTRACTION BEYOND THIS POINT
Received: May 01, 2012 Revised: May 02, 2012 Accepted: May 04, 2012