Hypothesis – Omega-6 to Omega-3 ratio increases obesity
Omega-3 and Omega-6 Fatty Acids: Role in Body Fat Gain and Development of Obesity
[N A JMedSci. 2015;8(4):157-165. DOI: 10.7156/najms.2015.0804157]
Lu Wang, MD, PhD lwang@research.bwh.harvard.edu
Division of Preventive Medicine, Brigham and Women’s Hospital, Boston MA
The ratio of m6 to m3 polyunsaturated fatty acid (PUFA) in western diet has remarkably increased over the past decades. This change in dietary fatty acid (FA) composition, independent of the total caloric intake and total fat intake, may contribute to the obesity epidemic in many populations. Experimental studies show that m3 and m6 FAs play different role in adipogenesis, lipid homeostasis, brain-gut-adipose axis signaling, and systemic inflammation, resulting in divergent effects on body fat growth. Evidence from human studies remains limited and inconclusive. Very few observational studies and clinical trials have examined the association between composition of PUFA, particularly m6 FA and rn6/m3 ratio, with obesity-specific parameters. A consensus on the optimal intake of m3 and m6 subtype FAs and m6/ra3 ratio in diet is lacking. We reviewed the temporal change in dietary PUFA composition in US, experimental studies that examine the effects of m3 and m6 FAs on body fat, and epidemiologic studies that assess the association between dietary PUFAs and the development of obesity. Future studies need to further evaluate dietary FAs and their biomarkers in association with objective and longitudinal measurements of body fat and elucidate the potential role of diet with a balanced m3 and m6 FA composition in the primary prevention of obesity.
Table 2
||Populations |Years| ω6/ω3 Ratio
Paleolithic| | 0.79
Greece |1960| 1.0-2.0
Japan |2000 |4.0
US |1909 |5.4
|1999| 9.6
France |1960 |4.1
|2000 |12
UK |2000 |15
India, rural| 2003 |5-6
India, urban| 2003 |38-50||
Figure F2
📄 Download the PDF from VitaminDWiki
INTRODUCTION
The ongoing obesity epidemic is a major public health issue in the US and worldwide. High fat intake has been implicated in the development of obesity,1,2 whereas evidence from prospective cohort studies 3-6 and randomized trials 7-10 linking total fat intake to body weight gain remains weak and inconsistent. In recent decades, total fat and saturated fat intake (as % of calories) in typical western diet has fallen 11,12 but the ratio of ra6 to m3 fatty acids (FAs) increased remarkably,13 in parallel with an alarming increase in the prevalence of overweight and obesity. Intake of m3 FAs, particularly long-chain m3 FAs that are mainly supplied by marine products, has shown benefits on multiple obesity- related disorders. ra6 FAs, more broadly available from vegetable oils to meat products, demonstrate less clear effect, with evidence suggesting possible harm.14 m3 and m6 FAs compete for common metabolic enzymes15 and incorporation to tissues.16 m3 and m6 FAs also may elicit opposing effects on adipogenesis,17 lipid homeostasis,18,19 brain-gut-adipose axis signaling,20 and systemic inflammation.21 Albeit strong laboratory evidence, epidemiologic data on the association between an imbalanced m3 and m6 FA intake with weight gain and obesity development is limited. With the lack of consensus on the optimal intake of m3 and m6 subtype polyunsaturated FA (PUFA) and the m6/m3 ratio, we reviewed experimental, population, and clinical study evidence regarding the effect of dietary PUFA composition (i.e. proportion of m3 and m6 FAs in total fat and the m6/m3 ratio) on the gain of body fat and development of obesity, independent of total caloric and total fat intake.
SIGNIFICANCE OF OBESITY EPIDEMIC AND OBESITY PREVENTION
Over the last few decades, the prevalence of overweight (defined as body mass index or BMI 25 to < 30 kg/m2) and obesity (BMI > 30 kg/m2) has dramatically increased in developed as well as developing countries.22-25 According to data from the National Center of Health Statistics (www.cdc.gov/nchs/data/factsheets), the prevalence of obesity has more than doubled since the 1970s among US adults aged > 20 years. Although recent estimates suggest that the overall rates of obesity have plateaued or even declined in some groups, the widespread obesity epidemic continues to be a leading public health problem in the US.26,27 Worldwide, nearly 500 million people 20 years and older were considered obese and 1.4 billion overweight in 2008.28 Obesity is a well-recognized modifiable risk factor for many adult-onset chronic diseases, such as cardiovascular disease (CVD),29 diabetes,30 hypertension,31 and cancer.32 The estimated total costs of health care for obesity-related conditions are as high as $139 billion annually.33,34 Effective strategies to control the obesity epidemic are urgently needed. Because the formation of adipose tissue appears to be an irreversible process,35 maintenance of normal body fat will have more efficient and sustainable health benefits than the treatment of established obesity,6,36 highlighting the importance of obesity primary prevention.
TEMPORAL CHANGE IN CONSUMPTION OF m3 AND o6 FA IN THE UNITED STATES
Since dietary fat represents a major source of energy, high fat i i tin oq intake has been considered a possible cause of obesity. , , However, evidence from prospective cohort studies3-6 and randomized trials7-10 linking total fat intake with gain of body weight or body fat is weak and inconsistent. This discrepancy may be explained by dietary fat composition.13,39 In the past half-century, the total fat and saturated fat intake (as % of calories) in industrial countries has continuously fallen,11,12 while the indiscriminate diet recommendations to substitute vegetable fats, which is high in m6 and low in m3 FAs, for animal fats have led to a substantial rise in intake of m6 FAs.13 These changes are accompanied by a significant increase in the supply of arachidonic acid (AA, 20:4m6, broadly available from vegetable oil to meat products) but no parallel increase in supply of long-chain m3 FAs (mainly from marine products) (Table 1).40 Moreover, substantial changes in animal feeds and food chain have been introduced.13 As a result, the ratio of m6 to m3 FAs in common Western diet has increased from the range of 1 to 4 41 to 10 to 40 (Table 2).13,40 This remarkable change in the balance of m3 and m6 FA intake may have contributed to the increasing prevalence of overweight and obesity in children and adults of many populations. In support of the recommendation by some research groups on reducing m6 FA intake to lower m6/m3 ratio,42,43 there is emerging evidence suggesting possible harm of excess m6 FA intake.14 However, the American Heart Association Scientific Advisory published in 2009 recommended m6 FA intake of at least 5-10% of energy, based on evidence that the consumption of m6 FAs (particularly linoleic acid, LA 18:2 m6) is associated with reduced risk of coronary heart disease.44 At the meantime, guidelines on specific intake of LA (18:2m6), a-linolenic acid (ALA, 18:3m3), and longer- chain PUFAs are formulated in several European countries by health authorities, but the recommended levels vary substantially.13 The optimal intake of m3 and m6 subtype PUFAs and m6/m3 ratio remains unresolved.
Table 1. Main Food Sources of m3 and m6 Fatty Acids in the United States in Year 1909 and 1999*.
- Originally from Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr. 2011;93:950-962. Data was obtained from the US Department of Agriculture (USDA).
Abbreviations: FA: fatty acid; AA: arachidonic acid (20:4 m6); LA: linoleic acid (18:2m6); ALA: a-linolenic acid (18:3 m3); EPA: eicosapentaenoic acid (20:5m3); DHA: docosahexaenoic acid (22:6m3); DPA: docosapentaenoic acid (22:5m3)
DIVERGENT EFFECTS OF m3 AND m6 FA ON DEVELOPMENT OF OBESITY
Advice to substitute PUFAs for saturated FAs (SFAs) is a cornerstone of worldwide dietary guideline since 1960s.45 By the time this guideline was initiated, PUFA was considered a uniform molecular category with beneficial effect of lowering blood lipids.46 Later research recognizes that PUFAs comprise of m3 and m6 subspecies, each having unique biochemical properties and playing different roles in metabolic outcomes including obesity. m3 and m6 FAs are distinguished based on the location of the first double bond. Both m3 and m6 FAs are essential for human because they cannot be synthesized and must be obtained from diet. Precursor FAs in both classes, ALA (18:3©3) and LA (18:2©6) respectively, can be desaturated and elongated to longer-chain FAs of the same class, albeit to a very low extent (Figure 1). In contrast, the two classes cannot be converted, and they are metabolically and physiologically distinct. In cultured cells, ©3 FAs inhibited,47 while ©6 FAs increased,48 cellular triglyceride content. In animal models, feeding mother rats with a high-fat diet rich in LA (18:2©6) resulted in hyperplasia or hypertrophy of white adipose tissue in suckling pups,49 whereas intake of fish oil rich in eicosapentaenoic acid (EPA, 20:5©3) and docosahexaenoic acid (DHA, 22:6©3) prevented the enhancement of fat mass.50,51 Thus, ©3 and ©6 FAs in diet potentially have divergent effects on the gain of body fat and the development of obesity.
POSSIBLE MECHANISMS OF DIVERGENT EFFECTS OF m3 AND m6 FA ON OBESITY DEVELOPMENT
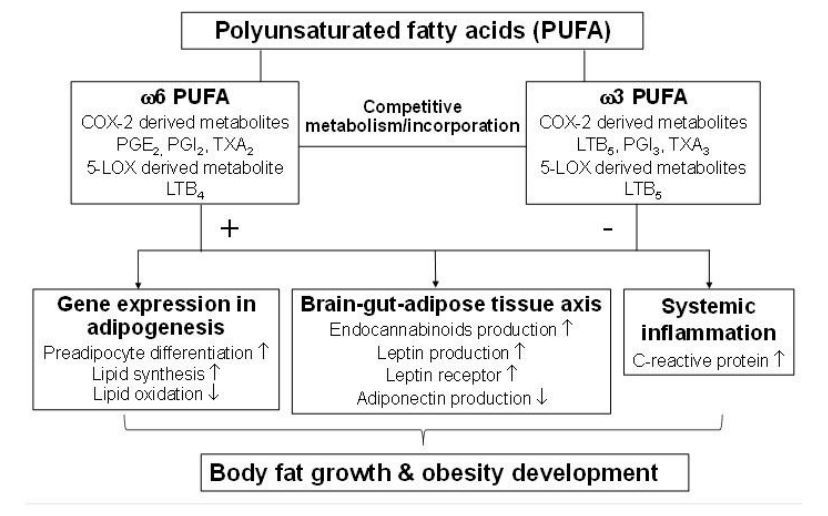
Due to the similarity in chemical structure, m3 and m6 FAs compete for common metabolic enzymes, both in the process of elongation and desaturation of precursor PUFAs and in the synthesis of downstream metabolites such as prostacycline and thromboxane.15 They also compete for incorporation into plasma lipid fractions and cell membranes.16 In addition to the competitions in metabolism, m3 and m6 FAs may have divergent effects on the development of obesity via several other mechanisms (Figure 2, Table 3).17-19
m3 AND m6 FA REGULATE GENE EXPRESSION IN ADIPOGENESIS AND LIPID METABOLISM
During the development of obesity, pluripotent stem cell precursors give rise to multipotent preadipocytes, and preadipocytes differentiate into mature adipocytes; lipid then fills in to cope with the high exogenous levels of fats.35 Because mature adipocytes do not divide in vivo or undergo significant turnover under physiological conditions, the proliferation of precursor cells and their differentiation into adipocytes are critical events in adipose tissue development.49 m3 and m6 FAs could serve as transcriptional factors to regulate the expression of genes involved in preadipocyte differentiation.17 m6 FAs such as AA (20:4m6) are potent adipogenic FA that up-regulates the expression of multiple adipogenic genes. In the early steps of preadipocyte differentiation, the metabolite of AA (20:4m6) by cyclooxygenase (COX) - prostacyclin - binds to its receptor (IP-R) on cell surface of preadipocytes, activates the protein kinase A (PKA) pathway,52 which ultimately up-regulates the expression of peroxisome proliferators-activated receptor (PPAR) family and leads to adipogenesis.53-57 The long-term adipogenic effect of AA is impaired by COX inhibitors.58 In IP-R knockout mice, no activation of PKA pathway occurs through IP-R, and thereafter the adipogenic effect of m6 FAs is similar to m3 FAs.58 In the later steps of preadipocyte differentiation, metabolites of AA (20:4m6) by lipoxygenases (LOX) also activate PPARs expression and promote adipogenesis.59
Table 3. Possible Mechanisms of Divergent Effects of m3 and m6 Fatty Acids on Obesity Development.
m3 FAs could inhibit the adipogenic effect of m6 FAs at multiple steps. First, m3 FAs such as DHA (22:6m3) have been shown to decrease the expression of COX-2 mRNA and production of COX-2 protein.60 Second, some m3 FAs, with potency of EPA(20:5m3) > DHA(22:6m3) > ALA(18:3m3), could inhibit the activities of COX-1 and COX-2.61 Third, long-chain m3 FAs could also directly inhibit the effect of prostaglandins arising from AA (20:4m6).62 Finally, m3 FAs [EPA(20:5m3) > DHA(22:6m3)] could down-regulate the production of cAMP49 or the catalytic subunit of PKA 63 in the prostacyclin-IP-R signaling pathway. In addition to these inhibitory effects on adipogenesis, m3 FAs also modulate lipid homeostasis by suppressing the expression of genes involved in lipogenesis including fatty acid synthase (FAS), lipoprotein lipase (LPL) and stearoyl-CoA desaturase-I (SCD-I) while increasing the expression of genes involved in P-oxidation such as acetyl-CoA oxidase (ACOX).18,19 The net result is a reduction in body fat deposition, mainly in liver and also in skeletal muscle and adipose tissue.
©3 AND ©6 FAs AND BRAIN-GUT-ADIPOSE AXIS SIGNALING
Another putative mechanism through which ©3 and ©6 FAs may affect body fat gain differently is the brain-gut-adipose axis. First, ©6 FAs derived metabolites endocannabinoids can mediate central control of appetite and energy balance. Endocannabinoids can be produced from the hydrolysis of AA-containing membranes.64 Receptors of endocanna binoids are present in all tissues involved in energy homeostasis.65 Activation of endocannabinoids receptors will stimulate food intake and lipogenesis in liver.66 ©3 FAs can reduce the endogeneous endocannabinoids production or lower the receptor sensitivity.67,68 Moreover, long-chain ©3 FA DHA (22:6©3) can induce another anorexigenic neuropeptide proopiomelanocortin in the hypothalamus, which curbs appetite and leads to weight loss.20
Second, ©3 and ©6 FAs may affect leptin signaling pathway differently. Leptin is a peptide hormone encoded by ob gene and is primarily produced and secreted by adipose tissue. The main function of leptin is the regulation of body weight by affecting appetite and energy expenditure.69 It has been postulated that human obesity might represent a state of leptin resistance.70 Dietary FA composition can affect leptin signaling. In rats, a diet rich in ©3 and ©6 FAs led to higher serum leptin levels than a diet rich in SFAs and monounsaturated FAs (MUFAs).71 When ©3 FAs were studied specifically, however, a high intake of ©3 FAs decreased the gene expression of leptin72 and leptin receptor73 both in vitro and in vivo. It is therefore possible that ©3 and ©6 FAs have divergent effects on leptin production, secretion as well as receptor signaling. In a small trial of healthy non-obese men (n = 30) and women (n = 25), the serum leptin concentration markedly decreased in women but not in men who received diet rich in MUFAs and ALA (18:3©3), despite only minor change in body weight and food intake. Serum leptin did not change in subjects who received diet rich in MUFAs only or ©6 FAs.74
Third, ©3 and ©6 FAs could modulate expression and secretion of adiponectin differently.75 Adiponectin is a plasma protein that is produced and secreted exclusively by adipocytes.76 Adiponectin suppresses a number of pathological processes linked with obesity. Experimental studies have shown that adiponectin activates 5’-AMP- activated protein kinase (AMPK),77 which stimulates FA oxidation and glucose utilization.78,79 These favorable effects ultimately lead to reduced lipid stores and improved insulin sensitivity in tissues.80 In mice model, diet rich in long-chain ©3 FAs EPA (20:5©3) and DHA (22:6©3) significantly increased the production and release of adiponectin in adipose tissue.81,82 One possible mechanism is that EPA (20:5m3) and DHA (22:6m3) activate PPARy, which in turn up-regulates the expression of adiponectin gene.82,83 In human studies, positive associations were found between circulating adiponectin and m3 FAs in plasma and adipose tissue.84,85 Diet intervention studies further demonstrated that long-chain m3 FA supplement raised plasma adiponectin concentration in human subjects.86 Few studies had examined the associations of m6 FAs with adiponectin. In a recent study of 44 end-stage renal disease patients, plasma adiponectin was positively associated with erythrocyte m3 FAs (r = 0.58) while inversely associated with m6 FAs (r = - 0.64) (both p < 0.01).87
m3 AND m6 FAs METABOLISM AND SYSTEMIC INFLAMMATION
The potential effects of m3 and m6 FAs are also related to the synthesis of specific eicosanoids through their downstream metabolism. The 2-series prostaglandins (PGE2, PGI2, and TXA2) and 4-series leukotrienes (LTB4) derived from m6 FAs by COX-2 and 5-LOX, respectively, are potent pro- inflammatory compounds. In contrast, 3-series prostaglandins (PGE3, PGI3, and TXA3) and 5-series leukotrienes (LTB5) derived from m3 FAs by the same enzymes are less pro-inflammatory. Since m3 and m6 FAs directly compete for access to the same converting enzymes, the balance in inflammatory status depends on the m6/m3 ratio. If m3 FA intake is high, m6 FA derived pro- inflammatory eicosanoids are inhibited, the risk of inflammatory conditions decreases. Additionally, the E series resolving (resolving E1 and E2, both formed by 5-LOX) derived from EPA (20:5m3) and the resolvins, protectins, and maresins derived from DHA (22:6m6) are all potent molecules that resolve inflammation, further enhancing the anti-inflammatory properties of m3 FAs. In animal models, m3 FAs, regardless provided in low-fat or high-fat diet, significantly lowered the expression of inflammatory markers such as monocyte chemoattractant protein-1 (MCP-1),88 interleukin-6 (IL-6),88 interferon-y (IFN-y),88 and plasminogen activator inhibitor type 1(PAI-1).89 In adipoctye-derived stem cells from the ob/ob mouse, EPA (20:5m3) and other m3 FAs significantly reduced IL-6 mRNA expression and secretion compared with AA (20:4m6) or LA (18:2m6).90 In human studies, m3 FAs rich diet or m3 FA supplementation significantly lowered m6/m3 ratio in plasma,91 circulating levels of C-reactive protein (CRP),92 IL- 6,91,92 tumor necrosis factor-a (TNF-a),93 MCP-1,94 and adipose tissue macrophage number.94 Laboratory and clinical studies specifically on m6 FAs or m6/m3 ratio with systemic inflammation are little. The direction of relation between obesity and inflammation has yet to be elucidated. It is well-accepted that accumulating adipose tissues promote the production of inflammatory cytokines and lead to an elevated chronic inflammation status. Alternatively, it is also possible that early stage of excess fat mass causes the change in growth factor environment over time, and results in a shift toward preadipocyte differentiation.95
EPIDEMIOLOGIC STUDIES OF PUFA INTAKE AND OBESITY
Distinctive associations of m3 and m6 FA intake with the development of several obesity-related disorders, such as hyperlipidemia and atherosclerotic disease, have been observed in population studies.41 Associations of m3 and m6 FA intake with the risk of type 2 diabetes, which is characterized by obesity and insulin resistance, are less clear. In prospective studies, the association between fish or EPA(20:5m3) + DHA(22:6m3) intake and incident type 2 diabetes has been mixed, with some studies in the US showing positive relations.96-98 There are fewer studies on m6 FAs: in cross-sectional studies, higher long-chain m6 FAs in skeletal muscle or erythrocyte phospholipids was associated with higher insulin sensitivity in healthy individuals,99 obese Pima Indians,100 and non-obese type 2 diabetic patients;101 while in prospective studies, higher y-linolenic acid (GLA, 18:3m6), eicosatrienoic acid (20:3m6), and AA (20:4m6) in serum cholesterol ester were associated with increased risk of type 2 diabetes.102,103
The associations of dietary PUFAs with the development of obesity per se have been rarely studied in human populations. We are aware of only two prospective cohort studies that examined dietary FAs in relation to longitudinal anthropometric change. Among 41,518 women in the Nurses’ Health Study, the intake of SFAs, MUFAs, PUFAs, and trans FAs was each weakly associated with weight gain during 8 years follow-up: the regression coefficients (P) on weight change (lbs) for each 1% difference in baseline FA intake (as % of calories) were 0.40, -0.31, 0.42, and 0.54, respectively (all P > 0.05).104 Among 16,587 men in the Health Professionals’ Follow-up Study, a 2% increment in energy intake from trans FAs that was isocalorically substituted for PUFAs was significantly associated with a 0.77 cm gain in abdominal circumference during 9 years follow-up (P < 0.001), and this association remained significant after controlling for concurrent change in BMI.105 These two studies have several limitations. First, both studies used self-administered semi-quantitative food frequency questionnaires (FFQ) to estimate dietary fat intake; second, both studies did not investigate m3 and m6 subtype PUFAs separately; third, both studies relied on self-reports to assess changes in anthropometry.
Our group recently conducted a prospective study in 534 participants of the Women’s Health Study (WHS) who had baseline measurement of erythrocyte FAs and a baseline BMI of 18.5 to < 25 kg/m2. Body weight was updated at a total of 6 time points during an average of 10.4 years follow-up. After multivariable adjustment for lifestyle and dietary factors including total energy intake and physical activity, the weight gain (kg) during follow-up in the highest versus the lowest quartile of baseline erythrocyte FAs was 3.08 vs. 2.32 for cis m6 FA (p trend: 0.04), 2.07 vs. 2.92 for cis m3 FA (p trend: 0.08), 2.93 vs. 2.05 for m6/m3 ratio (p trend: 0.046), and 3.03 vs. 2.27 for trans FA (p trend: 0.06).106 The Erythrocyte m6/m3 ratio was also positively associated with the risk of becoming overweight or obese (n = 186). To our knowledge, this is the first study that examined biomarkers of dietary PUFAs in association with longitudinal weight gain and newly developed obesity. However, the nature of this observational study did not allow investigation of the effect of fat composition change on adiposity measurement.
Several interventional studies have found that m3 FA supplementation can have beneficial effects on measurements of body weight and body fat in lean,107 overweight,108,109 and obese 110 individuals. Comparable study on m6 FA is lacking. We identified one diet intervention trial conducted in France that aimed to investigate the effect of an increased m3 FA combined with a reduced m6 FA consumption, with no change in total energy and other macronutrient intake.111 A total of 17 (10 men and 7 women) healthy, normal weight subjects with a low usual intake of m3 FAs were studied. After 10-week intervention via dietary recommendations focusing on m6/m3 ratio, intake of ALA (18:3m3) increased and LA (18:2m6) decreased, in erythrocyte membrane m3 FA increased while m6 FA did not change. The measurement of anthropometry including body weight, BMI, waist/hip ratio, fat mass, and plasma leptin concentration did not change significantly during the trial, but plasma adiponectin significantly increased (6.50 vs. 7.63 pg/ml at baseline and post-intervention, respectively) and plasma TNF-a (2.18 vs. 1.50 pg/ml), IL-6 (1.28 vs. 1.00 pg/ml), and CRP (0.23 vs. 0.17 mg/l) decreased. Major limitations of this trial include the small sample size, short-term intervention, and lack of a control group. There is clear need for more studies to further investigate the long-term effect of dietary m6/m3 ratio change and the optimal m6/m3 ratio for overall health.
CONCLUSION
m3 and m6 FAs are metabolically and physiologically distinct PUFA subclasses. Experimental studies suggest that m3 and m6 FAs may have divergent effect on body fat gain and development of obesity, though evidence on m6/m3 ratio specifically is little. Epidemiologic study of dietary PUFAs with the development of human obesity remains limited; there is currently no consensus on the optimal intake of m3 and m6 FAs and m6/m3 ratio. More studies are needed to examine the association of dietary m3 and m6 FAs, m6/m3 ratio, and their biomarkers with objective and longitudinal measures of obesity. These studies will provide insights regarding the long-term effects of dietary fat composition and whether a limit on m6 FA intake and maintenance of a balance between m3 and m6 FAs will promote a favorable change in body fat mass, curb epidemic of obesity, and prevent obesity-related health problems.
ABBREVIATIONS
FA: fatty acid; SFA: saturated fatty acid; MUFA: monounsaturated fatty acid; PUFA: polyunsaturated fatty acid; AA: arachidonic acid (20:4©6); LA: linoleic acid (18:2 w6); ALA: a-linolenic acid (18:3 w3); EPA: eicosapentaenoic acid (20:5©3); DHA: docosahexaenoic acid (22:6©3); DPA: docosapentaenoic acid (22:5©3); BMI: body mass index; CVD: cardiovascular disease; COX: cyclooxygenase; PKA: protein kinase A; PPAR: peroxisome proliferators-activated receptor; LOX: lipoxygenases; PG: prostaglandins; LT: leukotrienes; TX: thromboxane
REFERENCES
Hooper L, Abdelhamid A, Moore HJ, Douthwaite W, Skeaff CM, Summerbell CD. Effect of reducing total fat intake on body weight: systematic review and meta-analysis of randomised controlled trials and cohort studies. BMJ. 2012;345:e7666.
Tremblay A. Dietary fat and body weight set point. Nutr Rev. 2004;62:S75-77.
Carmichael HE, Swinburn BA, Wilson MR. Lower fat intake as a predictor of initial and sustained weight loss in obese subjects consuming an otherwise ad libitum diet. J Am Diet Assoc. 1998 Jan;98:35-39.
Heitmann BL, Lissner L, Sorensen TI, Bengtsson C. Dietary fat intake and weight gain in women genetically predisposed for obesity. Am J Clin Nutr. 1995;61:1213-1217.
Lissner L, Heitmann BL, Bengtsson C. Low-fat diets may prevent weight gain in sedentary women: prospective observations from the population study of women in Gothenburg, Sweden. Obes Res. 1997 Jan;5:43-8.
Sherwood NE, Jeffery RW, French SA, Hannan PJ, Murray DM. Predictors of weight gain in the Pound of Prevention study. Int J Obes Relat Metab Disord. 2000;24:395-403.
Sheppard L, Kristal AR, Kushi LH. Weight loss in women participating in a randomized trial of low-fat diets. Am J Clin Nutr. 1991;54:821-828.
Westerterp-Plantenga MS, Wijckmans-Duijsens NE, Verboeket-van de Venne WP, de Graaf K, van het Hof KH, Weststrate JA. Energy intake and body weight effects of six months reduced or full fat diets, as a function of dietary restraint. Int J Obes Relat Metab Disord. 1998;22:14-22.
Rock CL, Thomson C, Caan BJ, Flatt SW, Newman V, Ritenbaugh C, Marshall JR, Hollenbach KA, Stefanick ML, Pierce JP. Reduction in fat intake is not associated with weight loss in most women after breast cancer diagnosis: evidence from a randomized controlled trial. Cancer. 2001;91:25-34.
Brehm BJ, Spang SE, Lattin BL, Seeley RJ, Daniels SR, D'Alessio DA. The role of energy expenditure in the differential weight loss in obese women on low-fat and low-carbohydrate diets. J Clin Endocrinol Metab. 2005;90:1475-1482.
Troiano RP, Briefel RR, Carroll MD, Bialostosky K. Energy and fat intakes of children and adolescents in the united states: data from the national health and nutrition examination surveys. Am J Clin Nutr. 2000;72:1343S-1353S.
Swan G. Findings from the latest National Diet and Nutrition Survey. Proc Nutr Soc. 2004;63:505-512.
Ailhaud G, Massiera F, Weill P, Legrand P, Alessandri JM, Guesnet P. Temporal changes in dietary fats: role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog Lipid Res. 2006;45:203-236.
Ramsden CE, Zamora D, Leelarthaepin B, Majchrzak-Hong SF, Faurot KR, Suchindran CM, Ringel A, Davis JM, Hibbeln JR. Use of dietary linoleic acid for secondary prevention of coronary heart disease and death: evaluation of recovered data from the Sydney Diet Heart Study and updated meta-analysis. BMJ.346:e8707.
Simopoulos AP. Essential fatty acids in health and chronic disease. Am J Clin Nutr. 1999;70:560S-569S.
Fickova M, Hubert P, Cremel G, Leray C. Dietary (n-3) and (n-6) polyunsaturated fatty acids rapidly modify fatty acid composition and insulin effects in rat adipocytes. J Nutr. 1998;128:512-519.
Amri EZ, Ailhaud G, Grimaldi PA. Fatty acids as signal transducing molecules: involvement in the differentiation of preadipose to adipose cells. J Lipid Res. 1994;35:930-937.
Jump DB, Clarke SD, Thelen A, Liimatta M. Coordinate regulation of glycolytic and lipogenic gene expression by polyunsaturated fatty acids. J Lipid Res. 1994;35:1076-1084.
Clarke SD, Jump D. Polyunsaturated fatty acids regulate lipogenic and peroxisomal gene expression by independent mechanisms. Prostaglandins Leukot Essent Fatty Acids. 1997;57:65-69.
Schwinkendorf DR, Tsatsos NG, Gosnell BA, Mashek DG. Effects of central administration of distinct fatty acids on hypothalamic neuropeptide expression and energy metabolism. Int J Obes (Lond). 2011;35:336-344.
James MJ, Gibson RA, Cleland LG. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am J Clin Nutr. 2000;71:343S-348S.
Ogden CL, Yanovski SZ, Carroll MD, Flegal KM. The epidemiology of obesity. Gastroenterology. 2007;132:2087-2102.
Rennie KL, Jebb SA. Prevalence of obesity in Great Britain. Obes Rev. 2005;6:11-12.
Cameron AJ, Welborn TA, Zimmet PZ, Dunstan DW, Owen N, Salmon J, Dalton M, Jolley D, Shaw JE. Overweight and obesity in Australia: the 1999-2000 Australian Diabetes, Obesity and Lifestyle Study (AusDiab). Med J Aust. 2003;178:427-432.
Yumuk VD. Prevalence of obesity in Turkey. Obes Rev. 2005;6:9-10.
Flegal KM, Carroll MD, Kit BK, Ogden CL. Prevalence of obesity and trends in the distribution of body mass index among US adults, 19992010. JAMA. 2012;307:491-497.
Ogden CL, Carroll MD, Kit BK, Flegal KM. Prevalence of childhood and adult obesity in the United States, 2011-2012. JAMA. 2014;311:806-814.
WHO Media centre. Obesity and overweight Fact sheet N°311, May 2012. http://www.who.int/mediacentre/factsheets/fs311/en/ Accessed 10/30/2013.
Caterson ID, Hubbard V, Bray GA, Grunstein R, Hansen BC, Hong Y, Labarthe D, Seidell JC, Smith SC, Jr. Prevention Conference VII: Obesity, a worldwide epidemic related to heart disease and stroke: Group III: worldwide comorbidities of obesity. Circulation. 2004;110:e476-483.
Weinstein AR, Sesso HD. Joint effects of physical activity and body weight on diabetes and cardiovascular disease. Exerc Sport Sci Rev. 2006;34:10-15.
Huang Z, Willett WC, Manson JE, et al. Body weight, weight change, and risk for hypertension in women. Ann Intern Med. 1998;128:81-88.
Samanic C, Chow WH, Gridley G, Jarvholm B, Fraumeni JF Jr. Relation of body mass index to cancer risk in 362,552 Swedish men. Cancer Causes Control. 2006;17:901-909.
Finkelstein EA, Ruhm CJ, Kosa KM. Economic causes and consequences of obesity. Annu Rev Public Health. 2005;26:239-257.
Finkelstein EA, Fiebelkorn IC, Wang G. National medical spending attributable to overweight and obesity: how much, and who's paying? Health Aff (Millwood). 2003;Suppl Web Exclusives:W3-219-26.
Gregoire FM, Smas CM, Sul HS. Understanding adipocyte differentiation. Physiol Rev. 1998;78:783-809.
Katan MB. Weight-loss diets for the prevention and treatment of obesity. N Engl J Med. 2009;360:923-925.
Romieu I, Willett WC, Stampfer MJ, et al. Energy intake and other determinants of relative weight. Am J Clin Nutr. 1988;47:406-412.
Nguyen VT, Larson DE, Johnson RK, Goran MI. Fat intake and adiposity in children of lean and obese parents. Am J Clin Nutr. 1996;63:507-13.
Ailhaud G, Guesnet P, Cunnane SC. An emerging risk factor for obesity: does disequilibrium of polyunsaturated fatty acid metabolism contribute to excessive adipose tissue development? Br J Nutr. 2008;100:461-470.
Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr. 2011;93:950- 962.
Simopoulos AP. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp Bio Med (Marywood). 2008;233:674-688.
Simopoulos AP, Leaf A, Salem N, Jr. Essentiality of and recommended dietary intakes for omega-6 and omega-3 fatty acids. Ann Nutr Metab. 1999;43:127-30.
Hamazaki T, Okuyama H. The Japan Society for Lipid Nutrition recommends to reduce the intake of linoleic acid. A review and critique of the scientific evidence. World Rev Nutr Diet. 2003;92:109-132.
Harris WS, Mozaffarian D, Rimm E, et al. Omega-6 fatty acids and risk for cardiovascular disease: a science advisory from the American Heart Association Nutrition Subcommittee of the Council on Nutrition, Physical Activity, and Metabolism; Council on Cardiovascular Nursing; and Council on Epidemiology and Prevention. Circulation. 2009;119:902-907.
Dietary fat and its relation to heart attacks and strokes. Report by the Central Committee for Medical and Community Program of the American Heart Association. JAMA. 1961;175:389-391.
Keys A. Effects of Different Dietary Fats on Plasma-Lipid Levels. Lancet. 1965;1:318-319.
Strum-Odin R, Adkins-Finke B, Blake WL, Phinney SD, Clarke SD. Modification of fatty acid composition of membrane phospholipid in hepatocyte monolayer with n-3, n-6 and n-9 fatty acids and its relationship to triacylglycerol production. Biochim Biophys Acta. 1987;921:378-391.
Hennig B, Watkins BA. Linoleic acid and linolenic acid: effect on permeability properties of cultured endothelial cell monolayers. Am J Clin Nutr. 1989;49:301-305.
Massiera F, Saint-Marc P, Seydoux J, et al. Arachidonic acid and prostacyclin signaling promote adipose tissue development: a human health concern? J Lipid Res. 2003;44:271-279.
Parrish CC, Pathy DA, Angel A. Dietary fish oils limit adipose tissue hypertrophy in rats. Metabolism. 1990;39:217-219.
Raclot T, Groscolas R, Langin D, Ferre P. Site-specific regulation of gene expression by n-3 polyunsaturated fatty acids in rat white adipose tissues. J Lipid Res. 1997;38:1963-1972.
Vassaux G, Gaillard D, Ailhaud G, Negrel R. Prostacyclin is a specific effector of adipose cell differentiation. Its dual role as a cAMP- and Ca(2+)-elevating agent. J Biol Chem. 1992;267:11092-11097.
Barak Y, Nelson MC, Ong ES, et al. PPAR gamma is required for placental, cardiac, and adipose tissue development. Mol Cell. 1999;4:585-95.
Kubota N, Terauchi Y, Miki H, et al. PPAR gamma mediates high-fat diet-induced adipocyte hypertrophy and insulin resistance. Mol Cell. 1999;4:597-609.
Rosen ED, Hsu CH, Wang X, et al. C/EBPalpha induces adipogenesis through PPARgamma: a unified pathway. Genes Dev. 2002;16:22-26.
Ren D, Collingwood TN, Rebar EJ, Wolffe AP, Camp HS. PPARgamma knockdown by engineered transcription factors: exogenous PPARgamma2 but not PPARgamma1 reactivates adipogenesis. Genes Dev. 2002;16:27-32.
Rosen ED, Sarraf P, Troy AE, et al. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol Cell. 1999;4:611-617.
Gaillard D, Negrel R, Lagarde M, Ailhaud G. Requirement and role of arachidonic acid in the differentiation of pre-adipose cells. Biochem J. 1989;257:389-97.
Hihi AK, Michalik L, Wahli W. PPARs: transcriptional effectors of fatty acids and their derivatives. Cell Mol Life Sci. 2002;59:790-798.
Massaro M, Habib A, Lubrano L, et al. The omega-3 fatty acid docosahexaenoate attenuates endothelial cyclooxygenase-2 induction through both NADP(H) oxidase and PKC epsilon inhibition. Proc Natl Acad Sci U S A. 2006;103:15184-15189.
Ringbom T, Huss U, Stenholm A, Flock S, Skattebol L, Perera P, Bohlin L. Cox-2 inhibitory effects of naturally occurring and modified fatty acids. J Nat Prod. 2001;64:745-749.
Corey EJ, Shih C, Cashman JR. Docosahexaenoic acid is a strong inhibitor of prostaglandin but not leukotriene biosynthesis. Proc Natl Acad Sci U S A. 1983;80:3581-3584.
Mirnikjoo B, Brown SE, Kim HF, Marangell LB, Sweatt JD, Weeber EJ. Protein kinase inhibition by omega-3 fatty acids. J Biol Chem. 2001;276:10888-10896.
Silvestri C, Di Marzo V. The endocannabinoid system in energy homeostasis and the etiopathology of metabolic disorders. Cell Metab. 2013;17:475-490.
Di Marzo V. Endocannabinoids: synthesis and degradation. Rev Physiol Biochem Pharmacol. 2008;160:1-24.
Di Marzo V, Matias I. Endocannabinoid control of food intake and energy balance. Nat Neurosci. 2005;8:585-589.
Matias I, Bisogno T, Di Marzo V. Endogenous cannabinoids in the brain and peripheral tissues: regulation of their levels and control of food intake. Int J Obes (Lond). 2006;30 Suppl 1:S7-S12.
Batetta B, Griinari M, Carta G, et al. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese Zucker rats. J Nutr. 2009;139:1495-501.
Reidy SP, Weber J. Leptin: an essential regulator of lipid metabolism. Comp Biochem Physiol A Mol Integr Physiol. 2000;125:285-98.
Caro JF, Sinha MK, Kolaczynski JW, Zhang PL, Considine RV. Leptin: the tale of an obesity gene. Diabetes. 1996;45:1455-1462.
Cha MC, Jones PJ. Dietary fat type and energy restriction interactively influence plasma leptin concentration in rats. J Lipid Res. 1998;39:1655-1660.
Reseland JE, Haugen F, Hollung K, et al. Reduction of leptin gene expression by dietary polyunsaturated fatty acids. J Lipid Res. 2001;42:743-50.
Fan C, Liu X, Shen W, Deckelbaum RJ, Qi K. The Regulation of Leptin, Leptin Receptor and Pro-opiomelanocortin Expression by N-3 PUFAs in Diet-Induced Obese Mice Is Not Related to the Methylation of Their Promoters. Nutrition & metabolism. 2011;8:31.
Kratz M, von Eckardstein A, Fobker M, et al. The impact of dietary fat composition on serum leptin concentrations in healthy nonobese men and women. J Clin Endocrinol Metab. 2002;87:5008-5014.
Iwaki M, Matsuda M, Maeda N, et al. Induction of adiponectin, a fat- derived antidiabetic and antiatherogenic factor, by nuclear receptors. Diabetes. 2003;52:1655-1663.
Goldstein BJ, Scalia R. Adiponectin: A novel adipokine linking adipocytes and vascular function. J Clin Endocrinol Metab. 2004;89:2563-2568.
Arita Y, Kihara S, Ouchi N, et al. Paradoxical decrease of an adipose- specific protein, adiponectin, in obesity. Biochem Biophys Res Commun. 1999;257:79-83.
Hardie DG, Carling D, Carlson M. The AMP-activated/SNF1 protein kinase subfamily: metabolic sensors of the eukaryotic cell? Ann Rev Biochem. 1998;67:821-855.
Winder WW, Hardie DG. AMP-activated protein kinase, a metabolic master switch: possible roles in type 2 diabetes. Am J Physiol. 1999;277:E1-10.
Yamauchi T, Kamon J, Minokoshi Y, et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP- activated protein kinase. Nat Med. 2002;8:1288-1295.
Flachs P, Mohamed-Ali V, Horakova O, et al. Polyunsaturated fatty acids of marine origin induce adiponectin in mice fed a high-fat diet. Diabetologia. 2006;49:394-397.
Neschen S, Morino K, Rossbacher JC, et al. Fish oil regulates adiponectin secretion by a peroxisome proliferator-activated receptor- gamma-dependent mechanism in mice. Diabetes. 2006;55:924-928.
Banga A, Unal R, Tripathi P, et al. Adiponectin translation is increased by the PPARgamma agonists pioglitazone and omega-3 fatty acids. American journal of physiology Endocrinology and metabolism. 2009;296:E480-489.
Hernandez-Morante JJ, Milagro FI, Larque E, Lujan J, Martinez JA, Zamora S, Garaulet M. Relationship among adiponectin, adiponectin gene expression and fatty acids composition in morbidly obese patients. Obes Surg. 2007;17:516-524.
Fernandez-Real JM, Vendrell J, Ricart W. Circulating adiponectin and plasma fatty acid profile. Clin Chem. 2005;51:603-609.
Itoh M, Suganami T, Satoh N, et al. Increased adiponectin secretion by highly purified eicosapentaenoic acid in rodent models of obesity and human obese subjects. Arteriosclerosis, thrombosis, and vascular biology. 2007;27:1918-1925.
An WS, Son YK, Kim SE, et al. Association of adiponectin and leptin with serum lipids and erythrocyte omega-3 and omega-6 fatty acids in dialysis patients. Clinical nephrology. 2011;75:195-203.
Monk JM, Hou TY, Turk HF, et al. Dietary n-3 polyunsaturated fatty acids (PUFA) decrease obesity-associated Th17 cell-mediated inflammation during colitis. PLoS One. 2012;7:e49739.
Kalupahana NS, Claycombe K, Newman SJ, et al. Eicosapentaenoic acid prevents and reverses insulin resistance in high-fat diet-induced obese mice via modulation of adipose tissue inflammation. J Nutr. 2010;140:1915-1922.
Hsueh HW, Zhou Z, Whelan J, et al. Stearidonic and eicosapentaenoic acids inhibit interleukin-6 expression in ob/ob mouse adipose stem cells via Toll-like receptor-2-mediated pathways. J Nutr. 2011;141:1260-1266.
Itariu BK, Zeyda M, Hochbrugger EE, et al. Long-chain n-3 PUFAs reduce adipose tissue and systemic inflammation in severely obese nondiabetic patients: a randomized controlled trial. Am J Clin Nutr. 2012;96:1137-1149.
Tsitouras PD, Gucciardo F, Salbe AD, Heward C, Harman SM. High omega-3 fat intake improves insulin sensitivity and reduces CRP and
IL6, but does not affect other endocrine axes in healthy older adults. Horm Metab Res. 2008;40:199-205.
Zhao G, Etherton TD, Martin KR, Gillies PJ, West SG, Kris-Etherton PM. Dietary alpha-linolenic acid inhibits proinflammatory cytokine production by peripheral blood mononuclear cells in hypercholesterolemic subjects. Am J Clin Nutr. 2007;85:385-391.
Spencer M, Finlin BS, Unal R, et al. Omega-3 fatty acids reduce adipose tissue macrophages in human subjects with insulin resistance. Diabetes. 2013;62:1709-17.
Lepperdinger G. Inflammation and mesenchymal stem cell aging. Curr Opin Immunol. 2011;23:518-524.
Wu JH, Micha R, Imamura F, et al. Omega-3 fatty acids and incident type 2 diabetes: a systematic review and meta-analysis. Br J Nutr. 2012;107(Suppl 2):S214-227.
Wallin A, Di Giuseppe D, Orsini N, Patel PS, Forouhi NG, Wolk A. Fish consumption, dietary long-chain n-3 fatty acids, and risk of type 2 diabetes: systematic review and meta-analysis of prospective studies. Diabetes Care. 2012;35:918-929.
Xun P, He K. Fish Consumption and Incidence of Diabetes: metaanalysis of data from 438,000 individuals in 12 independent prospective cohorts with an average 11-year follow-up. Diabetes Care. 2012;35:930-938.
Borkman M, Storlien LH, Pan DA, Jenkins AB, Chisholm DJ, Campbell LV. The relation between insulin sensitivity and the fatty- acid composition of skeletal-muscle phospholipids. N Engl J Med. 1993;328:238-244.
Pan DA, Lillioja S, Milner MR, et al. Skeletal muscle membrane lipid composition is related to adiposity and insulin action. J Clin Invest. 1995;96:2802-2808.
Rodriguez Y, Christophe AB. Long-chain omega6 polyunsaturated fatty acids in erythrocyte phospholipids are associated with insulin resistance in non-obese type 2 diabetics. Clin Chim Acta. 2005;354:195-199.
Vessby B, Aro A, Skarfors E, Berglund L, Salminen I, Lithell H. The risk to develop NIDDM is related to the fatty acid composition of the serum cholesterol esters. Diabetes. 1994;43:1353-1357.
Wang L, Folsom AR, Zheng ZJ, Pankow JS, Eckfeldt JH, Investigators AS. Plasma fatty acid composition and incidence of diabetes in middle- aged adults: the Atherosclerosis Risk in Communities (ARIC) Study. Am J Clin Nutr. 2003;78:91-98.
Field AE, Willett WC, Lissner L, Colditz GA. Dietary fat and weight gain among women in the Nurses' Health Study. Obesity (Silver Spring). 2007;15:967-76.
Koh-Banerjee P, Chu NF, Spiegelman D, et al. Prospective study of the association of changes in dietary intake, physical activity, alcohol consumption, and smoking with 9-y gain in waist circumference among 16 587 US men. Am J Clin Nutr. 2003;78:719-727.
Wang L, Manson JE, Rautiainen S, et al. A prospective study of erythrocyte polyunsaturated fatty acid, weight gain, and risk of becoming overweight or obese in middle-aged and older women. European journal of nutrition. 2015.
Couet C, Delarue J, Ritz P, Antoine JM, Lamisse F. Effect of dietary fish oil on body fat mass and basal fat oxidation in healthy adults. Int J Obes Relat Metab Disord. 1997;21:637-643.
Krebs JD, Browning LM, McLean NK, et al. Additive benefits of long- chain n-3 polyunsaturated fatty acids and weight-loss in the management of cardiovascular disease risk in overweight hyperinsulinaemic women. Int J Obes (Lond). 2006;30:1535-1544.
Thorsdottir I, Tomasson H, Gunnarsdottir I, et al. Randomized trial of weight-loss-diets for young adults varying in fish and fish oil content. Int J Obes (Lond). 2007;31:1560-1566.
Kunesova M, Braunerova R, Hlavaty P, et al. The influence of n-3 polyunsaturated fatty acids and very low calorie diet during a shortterm weight reducing regimen on weight loss and serum fatty acid composition in severely obese women. Physiol Res. 2006;55:63-72.
Guebre-Egziabher F, Rabasa-Lhoret R, Bonnet F, et al. Nutritional intervention to reduce the n-6/n-3 fatty acid ratio increases adiponectin concentration and fatty acid oxidation in healthy subjects. Eur J Clin Nutr. 2008;62:1287-1293.