Vitamin K and bone – review
Clin Cases Miner Bone Metab. 2017 May-Aug;14(2):200-206. doi: 10.11138/ccmbm/2017.14.1.200. Epub 2017 Oct 25.
Maria Fusaro1’2 Maria Cristina Mereu3 Andrea Aghi4 Giorgio Iervasi1 Maurizio Gallieni5
📄 Download the PDF from VitaminDWiki
Where Vitamin K is stored (figure 2)
Summary
Vitamin K is mainly known as an agent involved in blood coagulation, maintaining the activity of coagulation factors in the liver. In addition, epidemiological studies suggested that a lack of vitamin K is associated with several diseases, including osteoporosis and vascular calcification. There are two main kinds of vitamin K: Phylloquinone (or PK) and Menaquinones (MKn), both act as co-enzyme of y-glutamyl carboxylase (GGCX) transforming under- carboxylated in carboxylated vitamin K dependent proteins, such as Bone Gla Protein (or Osteocalcin) and Matrix Gla Protein. Recently, Vitamin K was also identified as a ligand of the nuclear steroid and xenobiotic receptor (SXR) (in murine species Pregnane X Receptor: PXR), expressed in osteoblasts. The purpose of this literature review is to evaluate the protective role of Vitamin K in bone and vascular health.
Vitamin K is mainly known as an agent involved in blood coagulation, maintaining the activity of coagulation factors in the liver. In addition, epidemiological studies suggested that a lack of vitamin K is also associated with bone and mineral abnormalities, in particular osteoporosis, bone fractures, and vascular calcification.
Structure and source vitamin K
Vitamin K is the collective term for a family of fat-soluble compounds sharing a common 2-methyl-1,4-naphthoquinone ring, but differing in the side-chain at the 3-position (Figure 1). The three main forms are vitamin K1 or phylloquinone (PK), vitamin K2 or menaquinones (MKn) and vitamin K 3 or menadione. An isoprenoid side chain attached at the 3-position differentiates PK, which possesses a phytyl side chain, from MK form, the latter is characterized by a variable number of condensed isoprenoid units (MKn); while menadione is without side chain and it is a synthetic analog (1).
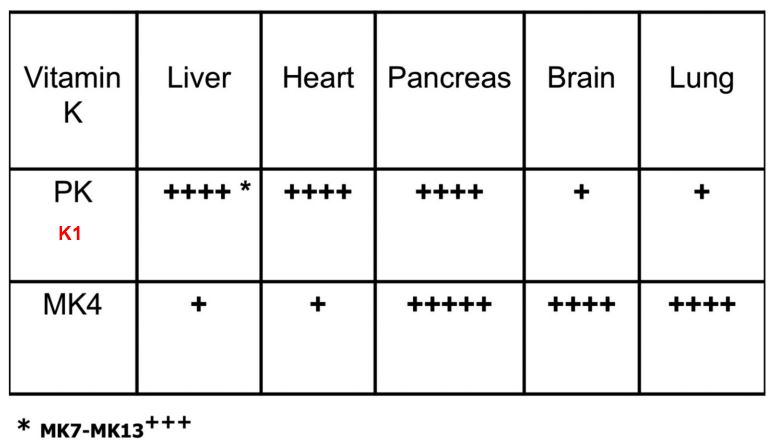
Figure 2 - Storage sites of vitamin K compounds. Vitamin K1 and MK-4 have different tissue distribution. Vitamin K1 is mainly present in the liver, heart and pancreas, while MK-4 is better represented in the kidneys, brain and pancreas. In the liver are also present long form K2: MK7 to MK13.
The sources of vitamin K are different depending on the vitamers. PK is found mostly in green leafy vegetable such as spinach, cauliflower and cabbage (2). In Western diets, MKn can be found in fermented foods such as butter, beef liver, curdled cheese, egg yolk. In Japan, natto is an important source: it is a traditional Japanese soybean-based food, primarily the product of fermentation using Bacillus Subtilis, and it is rich of Menaquinone-7 (MK-7) (2). Furthermore, MKn are also produced by the intestinal bacterial flora. Typically, these MKn have long lateral chains: MK-10 and MK-11 are synthesized by Bacteroides, MK-8 by Enterobacteria, MK-7 by Veillonella, and MK-6 by Eubacterium lentum (3, 4). The only exception is MK-4, which is not produced by the intestinal bacterial flora but it is converted from PK through a side chain removal/addition mechanism in specific tissues (pancreas, testes and vessel wall), with menadione as the intermediate molecule (5-7). Menadione, the synthetic form, is added in animal food and it is converted in the liver into MK-4 (8).
Metabolism and recycling of vitamin K
PK is absorbed by the small bowel with a saturable, energy- dependent mechanism (ATP), while MKn are absorbed in the colon by passive diffusion mechanism (1). Being fat-soluble, both forms of vitamin K need a normal pancreatic function and the presence of bile salts for their absorption (1). K vitamers are absorbed as mixed micelle into the enterocyte, assembling the chylomicrons in association with apolipoprotein A and B-48; ApoC and ApoE are added to the latter in the circulation. In the capillaries, chylomicrons are removed from triglycerides by lipoprotein lipase (LPL), also they become remnants, poor of triglycerides and rich of cholesterol, in the circulation. After reaching the liver, chylomicrons remnants are transformed into VLDL, which back into the circulation, together with Apolipoprotein B-100, C and E, enter into osteoblast cells as LDL interacting with lipoprotein receptors such as LDLR and LRP1. Thus, vitamin K enters into bone by endocytosis (1,9). Apoliporotein E is of particular interest, since Kohlmeier et al. have highlighted that vitamin K concentrations are markedly influenced by APO E polymorphism (ApO >2 Apo 3>Apo 4) (10).
MK-7 has a longer half-life than MK-4 and PK (2). Vitamin K is catabolized in the liver by a common degradative pathway, forming metabolites by oxidative degradation initially by w- hydroxylation followed p-oxidation pathway. Metabolites are excreted in the bile (about 40%) and in the urine (20%) as glucuronides (11).
Although vitamin K is fat soluble, only a little is stored in the body and without a regular dietary intake its reserve would be rapidly depleted. For this reason, the human body recycles Vitamin K, compensating its limited storage capacity (12). K vitamers storage is illustrated in Figure 2.
Vitamin K acts in the endoplasmic reticulum as coenzyme for the gamma-Glutamyl CarboXylase (GGCX) enzyme. The latter catalyzes the carboxylation of glutamic acid residues (Glu), which are transformed into gamma-carboxyglutamic acid (Gla), influencing the activity of Vitamin K-Dependent Proteins (VKDPs). Among the most relevant VKDPs are coagulation factors (phrothrombin, factor VII, IX, X, and proteins C, M, S, Z), Matrix Gla Protein (MGP), Gas6 (Growth Arrest-Specific 6 Protein), GRP (Gla Rich Protein) and Bone Gla Protein (BGP, or osteocalcin) (13-16).
Warfarin acts as a Vitamin K antagonist by blocking two important reactions of the vitamin K cycle: vitamin K epoxide reductase (VKOR) and vitamin K quinone reductase, resulting in a consequent functional vitamin K deficiency (17) (Figure 3). Vitamin K epoxide reductase complex subunit 1(VKORC1) can be a marker for bone disease. Its polymorphisms have been associated with BMD alterations (both lower and higher) (18, 19). Recently, Vermeer et al. (20) synthesized novel 2-methyl-1,4-naphthoquinone derivatives, differing in the side-chain at the 3-position. They demonstrated their biological activity as carboxylase agonists compounds, in particular they found that compounds 13 (C-16 tail) and 15 (C-14 tail) showed strong bioactivity while compounds 12 (C-17 tail) and 14 (C-18 tail) showed vice versa a weak activity. This finding is particularly interesting, because compared to naturally occurring forms of vitamin K, these synthetic vitamin K derivatives with higher bioactivity might be used in pharmacological interventions, rather than using traditional vitamin K supplements, in the clinical setting (20).
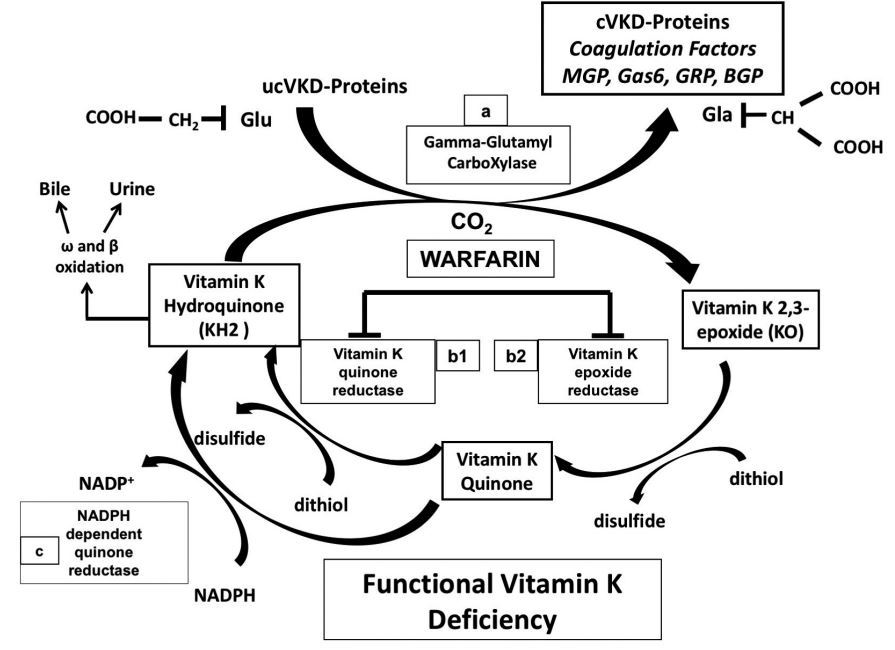
Figure 3 - The vitamin K cycle. Vitamin K acts as a coenzyme.
The enzyme involved in this reaction is Gamma-Glutamyl-CarboXylase (GGCX) (a).
The catalyzed reaction is the carboxylation of glutamic acid residues (Glu) which are transformed into gamma-carboxyglutamic acid (Gla).
Warfarin acts as a vitamin K antagonist by blocking two important reactions: vitamin K quinone reductase (b1) and Vitamin K epoxide reductase (VKOR) (b2), resulting in a consequent functional vitamin K deficiency. NADPH dependent quinone reductase (c) is relatively insensitive to the inhibiting action of warfarin and represent an escape pathway for the transformation of vitamin K quinone into vitamin K hydroquinone. MGP: Matrix Gla Protein, Gas6: Growth arrest-specific 6, GRC: Gla-rich protein, BGP: Bone Gla Protein.
The vitamin K receptor
Recently a different mechanism of action has been highlighted for vitamin K. In fact, it may act as ligand of the nuclear receptor, Steroid and Xenobiotic Receptor (SXR) (Pregnane X Receptor, PXR, murine homolog). SXR is expressed in osteoblastic cell lines and it is activated by vitamin K. In particular, SXR/PXR forms heterodimers with the 9-cis-retinoid acid receptor (RXR) and this latter complex binds to SXR-re- sponsive elements (SXRE) within the promoter or enhancer regions of target genes (21). The activation of SXR/PXR in bone tissue could mediate the beneficial effects of vitamin K. Accordingly, Azuma et al. (21) identified genes induced by vitamin K in an SXR-dependent manner, including tsukushi (Tsk), matrilin-2 (Matn2) and CD14. Tsk encodes a protein that has a collagen-accumulating effect, Matn2 is a protein comprising an extracellular matrix like collagen, whereas CD14 regulates osteoblastogenesis and osteoclastogenesis by inducing differentiation of B cells (21). They also found that 4-month-old female PXR-deficient mice showed lower bone mineral density in femoral bone. Micro Computed Tomography images of the femoral trabecular bone showed a fragile structure with bone loss in PXR-deficient mice (22). Induction of these genes is not inhibited by warfarin treatment, pointing to a GGCX-independent mechanism. Thus, SXR/PXR promotes bone formation and suppresses bone resorption, indicating that SXR/PXR may be a key regulator of bone homeostasis.
In addition, MKn seem to inhibit osteoclastic bone resorption, by suppression of RANKL expression (23, 24).
Vitamin K status and vitamin K dependent proteins activity: osteocalcin/Bone Gla Protein (BGP)
Vitamin K status can be measured indirectly by a VKDP activity parameter, such as the prothrombin time, or by measurement of under-carboxylated proteins. The most relevant proteins are under-carboxylated prothrombin or PIVKA-II (Protein Induced by Vitamin K Absence/Antagonism II, threshold: > 2 nmol/L); under-carboxylated osteocalcin (or ucBGP, threshold: >20%); under-carboxylated mGp (ucMGP) and/or de-phosphorylated under-carboxylated MGP (dp-ucMGP) (25).
The amount of under-carboxylated BGP could represent a more sensitive marker of vitamin K status in humans, better detecting subclinical vitamin K deficiency than prothrombin time (26).
Few published studies deal with the direct measurement of K vitamers. Most are focused on PK levels, starting from the study of Sadowski et al. (27), who showed that although the elderly subjects had higher absolute levels of phylloquinone than the young subjects (means = 1.05 vs 0.94 nmol/L), when the concentration of phylloquinone in plasma was expressed as nmol phylloquinone per mmol of triglycerides, the elderly subjects had significantly decreased levels for phyllo- quinone compared with the young subjects (0.62 vs 0.82 x 10(-6). These findings were later confirmed in hemodialysis patients by Pilkey et al. (PK levels <0.4 nM/L) (28) and in our multicenter VIKI study (29).
Recommendations for daily intake of vitamin K are inconsistent. The Institute of Medicine (US) Panel on Micronutrients proposed an adequate intake for men and women of 120 and 90 ug/day, respectively (30). In 2012, the Italian LARN (Reference Assumption Levels for Nutrients and Energy), proposed by the Human Nutrition Italian Society (SINU), suggested an intake of vitamin K stratified for age (140 or 170 ug/day for 18-59 and >60 years old, respectively) (31).
The predominant dietary form of vitamin K in the USA, Europe, and most Western countries is phylloquinone, while the major form in Japan is menaquinones, especially menaquinone 7 (MK-7), which is a component of natto (3). Few data are available on vitamin K intake both in the general population and in chronic kidney disease patients (32). Recently, we evaluated the dietary intake of PK in a cohort of patients undergoing hemodialysis (33). We measured vitamin K1 intake in 91 hemodialysis patients (median age 71 years) compared to 85 healthy adults (69 years). Participants completed a food journal of seven consecutive days for the estimation of dietary intakes of macro- and micro-nutrients. The average total daily vitamin K1 intake was 72 ug/day in dialysis patients, compared to 129 iug/day in healthy controls, indicating a low PK intake in hemodialysis patients undergoing a Mediterranean diet, compared to control subjects with normal renal function (33).
Osteocalcin/BGP is a small protein of 49 aminoacids, containing three Gla residues. It is produced by osteoblasts and odontoblasts under the control of vitamin D, a known promoter for BGP gene synthesis. Active BGP (carboxylated:cBGP) is able to bind calcium ions and incorporate them in the hydroxyapatite crystals that form the bone matrix (34,35). cBGP and under-carboxylated (ucBGP) concentrations reflect the functional status of this protein in the bone matrix and they have been shown to be a good index to describe bone health (36).
Warfarin treated male patients have a higher vertebral fracture risk, both in normal renal function (37) and CKD patients (38). In the latter population, we found an association between vertebral fractures rates and lower total BGP levels and/or higher ucBGP especially in males (38). Moreover, recent findings support an indirect role of BGP in the protection against vascular calcifications, through several mechanisms, including extraskeletal effects on glucose and lipid metabolism (39, 40). Another possible mechanism of protection from vascular calcifications can be mediated by adiponectin, which can be increased by BGP secretion in rats with chronic kidney disease (41). Adiponectin, an anti-inflammatory adipokine, is secreted by adipocytes and in the arterial wall, preventing the transdifferentiation of vascular smooth muscle cells into osteoblast-like cells (42, 43).
Vitamin D and Vitamin K
Vitamins D and K have osteoinduction properties (44). Koshihara et al. highlighted that MKn enhances vitamin D3 induced mineralization, possibly through accumulation of cBGP in the extracellular matrix as well as increased BGP gene expression (45).
Gigante et al. (46) showed that supplementation with vitamins MK-7, D3 and a combination of both was able to modulate the expression of genes involved in both mineralization and angiogenesis, and that Vitamin MK-7 enhances vitamin D3 effects on osteoblast precursors mesenchymal stem cells. The study reveals a novel and potent effect of vitamin D3 in association with vitamin MK-7 in inducing bone development by mesenchymal stem cells. Further studies in vivo should be conducted to verify how these molecular effects translate into accelerated bone healing (46).
Vitamin K deficiency and fractures
Several studies suggest that vitamin K deficiency is related to osteoporosis, pathological fractures and vascular calcifications. Low intake of PK has recently been associated with an increase in the risk of hip fracture in the general population, and treatment with vitamin K may reduce the relative risk for non vertebral and hip fractures (47, 48). Moreover, consumption of natto, a Japanese food rich in vitamin K, is associated with significantly higher bone mineral density (BMD) in elderly Japanese men, and high dietary intake of vitamin K is associated with higher BMD in elderly subjects of both genders (49). Conversely, low dietary vitamin K intake is associated with low BMD in women of all ages (50).
Chronic kidney disease-related bone and mineral disease (CKD-MBD) is defined as a systemic disorder of mineral and bone metabolism due to CKD, which is manifested by abnormalities in bone and mineral metabolism and/or extraskeletal calcification (51). Although CKD-MBD is one of the possible causes of osteoporosis, we do not have evidence that CKD- MBD is the only or crucial determinant of bone mechanical strength in CKD patients (52). The risk of hip fracture is considerably high in CKD patients. Tentori et al. (53) evaluated the incidence of hip fracture among participants of the Dialysis Outcomes and Practice Patterns Study (DOPPS); they showed a variable incidence of hip fractures across countries, from 12 events/1000 patient-years in Japan to 45/1000 patient-years in Belgium. In all countries, fracture rates were higher in hemodialysis patients compared to those reported for the general population. Only the in Japanese people fracture rates were similar for both groups (53). Possibly, the high dietary intake of vitamin K in Japan could in part be associated to this finding.
We carried out an observational study (29), where we assessed in 387 hemodialysis patients PK and MKn (MK-4, MK-5, MK-6 and MK-7, adjusted for triglycerides) levels and their association with vertebral fractures by quantitative vertebral morphometry and aortic and iliac calcifications with the same X-ray of spine (D5-L4). We found deficiency of MK-7 (35.4%), PK (23.5%), and MK-4 (14.5%). A total of 55.3% of patients had vertebral fractures, 80.6% had aortic calcifications and 56.1% had iliac calcification. PK deficiency was the strongest predictor of vertebral fractures (OR 2.94), MK-4 deficiency was the strongest predictor of aortic calcification (OR 2.82) (29). Moreover, in the same study we found lower BGP levels in patients with high prevalence of vertebral fractures and vascular calcifications.
Warfarin use may predispose to bone fractures and vascular calcification by different mechanisms :
directly, by inhibition of y-carboxylation of BGP and
indirectly, because patients treated with warfarin may limit their dietary intake of foods rich in vitamin K.
New oral anticoagulant seems to have less influence on bone metabolism (54), but their long-term effects need more studies. An experimental study in normal rats treated with dabigatran vs warfarin highlighted the bone response to anticoagulant treatment. Bone was evaluated immuno-histochemically and histomorphometrically: dabiga- tran treated animals preserved femur and vertebral volume and structure, with higher bone volume and trabecular thickness and less trabecular separation, compared to rats treated with warfarin. The thinner trabeculae with increased separation in the marrow space of warfarin treated animals suggested a potentially increased fragility, despite similar microarchitecture (54). This finding is consistent with clinical observations from the American Atrial Fibrillation registry (37), where males treated with warfarin over one year had a higher prevalence of vertebral fractures than controls. Similar results were described in the dialysis population, associated with lower BGP levels in males (38).
Vitamin K as therapeutic tool for the prevention of fractures
Only a few randomized controlled studies evaluated the impact of vitamin K supplementation on fractures. A key finding of these studies is that vitamin K supplementation at high doses has a positive effect on the skeleton of postmenopausal women with a reduced incidence of fracture mediated by mechanisms other than bone mineral density increase and bone turnover control (55). A systematic review (56) published in 2006 suggested that supplementation with PK and MK-4 reduces bone loss. Moreover, it indicated for MK-4 a strong effect on incident fractures among Japanese patients. From a clinical perspective, the results of this review suggested that patients at risk for fracture should be encouraged to consume a diet rich in vitamin K but, on the other hand, the Authors suggested that routine supplementation is not justified until these results are confirmed in a large pragmatic study with fractures as the main outcome (56). Still, the available studies are pointing to a relevant effect of high dose vitamin K treatment on bone fractures. Shiraki et al. (57) studied 241 osteoporotic patients in a 24-month randomized open label study. The vitamin K2-treated group received MK-4 (menatetrenone) 45 mg/day orally. Patients were followed for lumbar bone mineral density and occurrence of new clinical fractures. As expected, treatment enhanced BGP y-carboxylation, while markers of bone turnover were unchanged. High dose MK-4 treatment significantly reduced the occurrence of new bone fractures (15% of patients affected by fractures, mainly vertebral, compared to 35% in the control group) and maintained lumbar bone mineral density (57).
Vitamin K1 also proved to be effective in a randomized trial. Cheung et al. (58) administered daily PK 5 mg or placebo in 440 postmenopausal women with osteopenia for 2 years in a randomized fashion. Despite a lack of effect on bone mineral density and bone resorption, the incidence of clinical vertebral fractures among women in the PK group was significantly lower (9 versus 20, p < 0.05). They suggested that the effect of PK on bone may not be related to changes in bone mineral density or bone turnover, but rather on PK effects on bone quality. Of interest, patients participating in this study were vitamin D replete, with a mean serum 25-hydroxyvita- min D level of 77 nmol/l at baseline. The issue of the effects of both vitamin K and vitamin D on hip fractures was addressed in a recent case-control study. Torbergsen et al. (59) found that serum concentrations of vitamin K1 and 25(OH)-vitamin-D were lower and significantly related to the risk of fracture in aged patients admitted for hip fracture. In addition to their independent roles, there was a synergistic effect between vitamin K1 and 25(OH)-vitamin-D.
Conclusions
Both PK and MKn may provide significant improvement in BGP levels and a reduced incidence of spine fractures. The most relevant effects have been observed with high doses of MK-4, up to 45 mg/day, which appear to be safe. Vitamin K1 supplementation appears to mostly affect bone quality, rather than bone mineral density. However, MK-4 may have a positive impact on BMD as well. Vitamin D status should be taken into account in future studies on vitamin K effects on bone, as there might be an interaction between the two molecules. Treating osteoporotic patients with vitamin K might have the additional advantage of protecting arteries from vascular calcification through its action on MGP, a vitamin K dependent protein with anti-calcific properties. This might prove particularly important in patients with chronic kidney disease, where other treatments for osteoporosis, such as bisphosphonates, cannot be used. Studies on vitamin K supplementation in CKD patients are ongoing, mainly aimed at the surrogate but robust outcome of vascular calcifications, and might provide further information on the best therapeutic approach.