Nutrients, Microbiome, and Chronic Diseases
The microbiome as a major function of the gastrointestinal tract and its implication in micronutrient metabolism and chronic diseases
Nutr Res . 2023 Mar 4;112:30-45. doi: 10.1016/j.nutres.2023.02.007
Dingbo Lin 1, Denis M Medeiros 2
PDF Table of Contents
2 Small intestine in nutrient digestion and absorption
3 Large intestine
4 Overview of the intestinal microbiome
5 Vitamin C and other redox-active food compounds on the gut microbiome
6 Gut microbiome-derived folate does not meet human needs
7 Carotenoids' interaction with the microbiome
8 Vitamin A: more than meets the eye
9 Vitamin D: a hormone communicating with the microbiome
10 Zinc: an essential nutrient for the gut microbiome and the host immunity
11 Iron and the microbiome
12 Limitations of current studies
13 The concept of precision microbiome
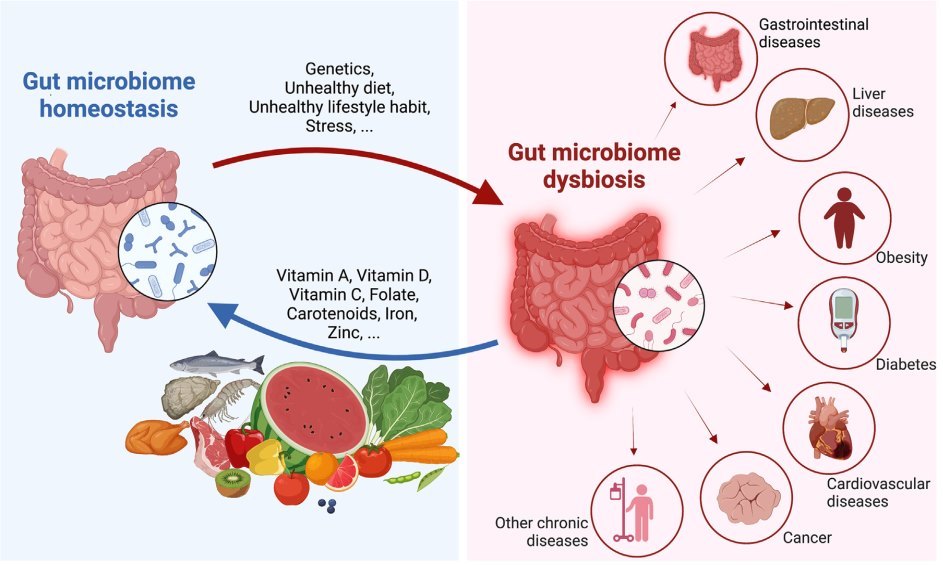
The composition and function of microbes harbored in the human gastrointestinal lumen have been underestimated for centuries because of the underdevelopment of nucleotide sequencing techniques and the lack of humanized gnotobiotic models. Now, we appreciate that the gut microbiome is an integral part of the human body and exerts considerable roles in host health and diseases. Dietary factors can induce changes in the microbial community composition, metabolism, and function, thereby altering the host immune response, and consequently, may influence disease risks. An imbalance of gut microbiome homeostasis (i.e., dysbiosis) has been linked to several chronic diseases, such as inflammatory bowel diseases, obesity, and diabetes. Remarkable progress has recently been made in better understanding the extent to which the influence of the diet-microbiota interaction on host health outcomes in both animal models and human participants. However, the exact causality of the gut microbiome on the development of diseases is still controversial. In this review, we will briefly describe the general structure and function of the intestine and the process of nutrient absorption in humans. This is followed by a summarization of the recent updates on interactions between gut microbiota and individual micronutrients, including carotenoids, vitamin A, vitamin D, vitamin C, folate, iron, and zinc. In the opinion of the authors, these nutrients were identified as representative of vitamins and minerals with sufficient research on their roles in the microbiome. The host responses to the gut microbiome will also be discussed. Future direction in microbiome research, for example, precision microbiome, will be proposed.
📄 Download the PDF from VitaminDWiki
Section 9 Vitamin D: a hormone communicating with the microbiome
Vitamin D is 1 of the most important vitamins because it also has a role as a hormone, thus allowing a greater number of reactions it is responsible for. This vitamin is found in bacteria, fungi, protozoans, and viruses [7]. A lack of vitamin D can lead to inflammatory diseases, such as IBS, Crohn disease, and ulcerative colitis. These diseases interact with genes, environment, and gut microflora [85,86]. Dietary supplementation of vitamin D is associated with the mitigation of IBS [87]. This vitamin requires a nuclear receptor as a partner and, specifically, the gut epithelium is restored to normal in the host by the vitamin D/nuclear receptor complex. This suggests the complex may inhibit inflammation in IBS [88,89]. The microbiome has these receptors that influence xenobiotic metabolism, fat storage, and renewal of GI epithelial cells [88,89].
It appears that the roles of vitamin D influence the immune system as an immune mediator [89]. Vitamin D deficiency is more common in patients with IBS compared with healthy populations. The active form of vitamin D (1,25-(OH)2 - vitamin D3 ) regulates the GI microbiome and promotes anti inflammatory and tolerogenic immune responses [89]. Innate immunity as a result of vitamin D3 induces antimicrobial compounds by promoting T cells and cytokines in the host [90].
An unanswered question is “How does deletion of vitamin D affect the microbiota metabolites?” [89,91]. Mice with tissue- specific vitamin D receptor (VDR) deletions that are tissue- specific in intestinal epithelial cells and myeloid cells were used to answer this question [92]. Microbiota metabolites from the host carbohydrates, proteins, lipids, and bile acids were changed in mice with the deletion of VDR [91]. Deletion of VDR affected 84 of more than 700 biochemicals because VDR status compared with 530 changes resulting from a high-fat diet [91].
Multiple sclerosis is a neuroinflammatory disease. The progression of this disease can be modulated by the gut microbiota, diet, and vitamins A and D [93]. The mechanisms of how the microbiome impacts vitamin D metabolism and multiple sclerosis are mixed. Microbiota-derived metabolites can also regulate VDR gene expression in the host, thus indirectly impacting vitamin D metabolism and function [93]. A high saturated-fat diet induces gut microbiome compositional shifts and functional changes, and subsequential intestinal inflammation, thereby altering the blood-brain barrier and resulting in neuroinflammation [94].
A vitamin D deficiency may lead to increased colorectal cancer risk. A case-control study of human fecal microbiota with vitamin D supplementation was conducted. The data were adjusted for vitamin D and other risk factors and revealed that the microbiome with an increased Bifidobacteria/Escherichia ratio is thought to decrease colorectal cancer risk [95].
Tabatabaeizadeh et al. reported compositional changes in the microbiome of adolescent girls given vitamin D supplements, Bacteroidetes and Lactobacillus decreased with vitamin D supplementation, whereas Firmicutes and Bifidobacterium increased [96]. This demonstrates the impact that a high vitamin D intake can change the composition of the gut microbiome in humans. Future studies are needed for a better understanding of the mechanisms and the benefits achieved by which vitamin D affects the gut microbiome [96].
Section 10 Zinc: an essential nutrient for the gut microbiome and the host immunity
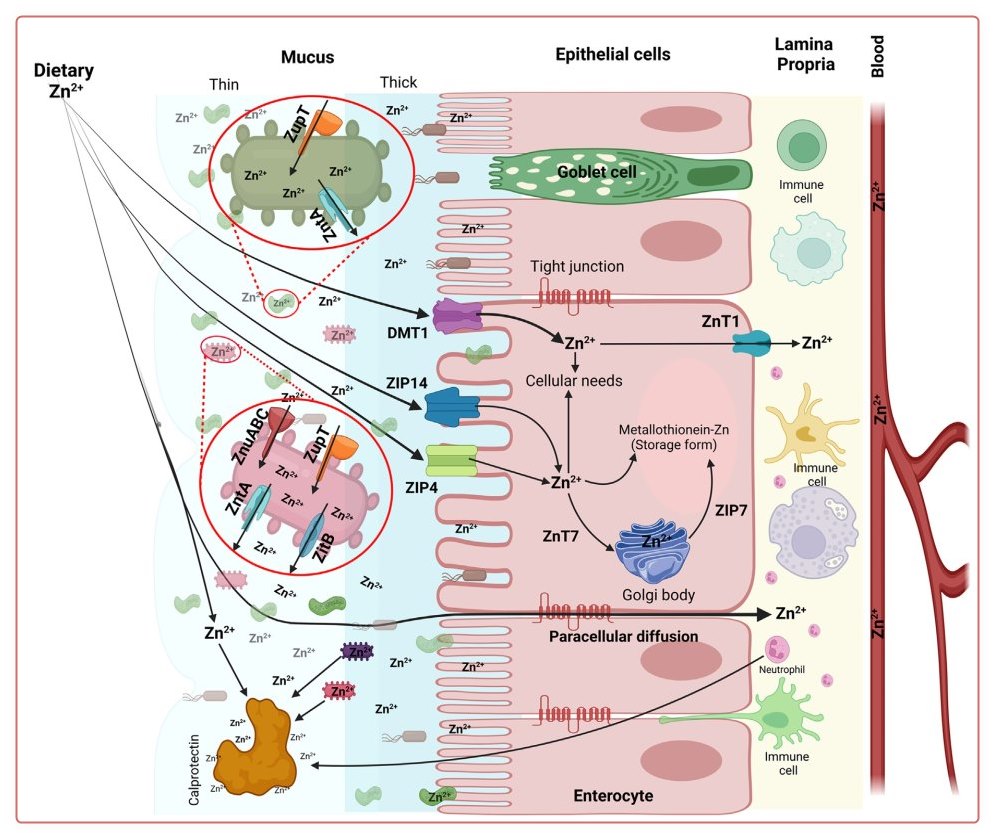

Zinc is a trace element and exerts many functions in host cells and the gut microbiome (Fig. 3). In the host GI tract, dietary zinc is liberated to the free form from dietary protein before absorption in the enterocyte and gut microbes. At physiologic levels of intestinal zinc, the predominant mechanism of absorption appears to be transporter-mediated via divalent metal transporter 1, zinc transporter 4 (ZIP4), and ZIP14 [97-103]. Zinc also diffuses through the intestinal mucosa into the lamina propria by paracellular diffusion [7]. Absorbed zinc is exported out of the enterocyte by Zn transporter 1 (ZnT1) [98-101]. ZnT7 and ZIP7 are carriers in the intracellular transport of zinc. Metallothionein (MT) is a zincbinding protein to store zinc as an intracellular buffering pool in the mucosa and other types of cells (Fig. 3). Zinc supplementation in humans can decrease inflammation of the GI tract. This observation may lead to an increase in MT levels, followed by a decrease in inflammation [99,104106]. On the other hand, low MT expression is present in inflammatory bowel disease patients [106] and intestinal colitis rodents [107].
Zinc is an essential trace nutrient for gut microbiota as well, though the information on the metabolic mechanism is still limited. Therefore, as shown in Table 2, a list of several zinc-dependent enzymes and their functions in mammalian cells would be beneficial for understanding zinc metabolism in gut microbiota. Presumably, there would be some similar or comparable metabolic features in these microbes, compared with mammals. Zinc homeostasis in gut microbes is maintained by zinc uptake via ZnuABC and ZupT transporters and zinc export by exporters ZntA and ZitB (Fig. 3) [104,105,108,109]. For some Enterobacteriaceae with limited zinc availability from the environment (as being sequestrated by calprotectin) and/or the deletion of zinc transporters, their colonization in the inflamed gut is enhanced via a zincophore- mediated zinc acquisition mechanism in zinc-limited media [110].
The studies demonstrated that zinc could affect gut micro- biome composition and function [104]. Abnormal gut physiology and microbiota composition may result in an increase in inflammatory proteins during pregnancy [111]. On the other hand, microbiota composition, gut pathology, and cytokines can be rescued by an amino acid-conjugated zinc complex, suggesting that a zinc deficiency may contribute to abnormal gut-brain signaling by altered physiology and microbiota composition [112]. In school-age children, zinc deficiency is associated with the elevation of serum inflammatory cytokines, an increase in the Shannon index (an indicator of alpha diversity of microbial communities) of the fecal microbiome, and enhanced richness in fecal Coprobac- ter, Acetivibrio, Paraprevotella, and Clostridium_XI [113]. In contrast, increased zinc in the gut results in the alteration of gut microbiota that appears linked to inflammation and pancreatitis, autism spectrum disorder, attention deficit disorder, attention deficit hyperactivity disorder, and fetal alcohol syndrome [114].
It appears that there is a zinc competition among the microbiota population in the GI tract. Zinc uptake by a high- affinity ZnuABC transporter is required for Campylobacter jejuni survival and growth in chickens with a normal microbiota but not when chickens are raised with a limited microbiota [115]. Without the ZnuABC transporter, C jejuni is unable to replicate or colonize in the GI tract [115], implicating that limiting zinc levels in chicken could limit human infection caused by C jejuni.
In addition to the deleterious changes mentioned earlier in zinc deficiency, autism spectrum disorder appears limited to gut problems and altered microbiota. Mice made zinc deficient in the prenatal stage appear to have autism spectrum behavior [111]. Biological studies collectively exhibit altered gut physiology and proinflammatory signaling. Neuroinflammation and gut composition are similar to autism spectrum disorder in humans [111].
Excess diet zinc can decrease the level of antibiotics to confer susceptibility to Clostridium difficile [116]. The Zn-binding S100 protein calprotectin shows antimicrobial effects against C difficile through chelating zinc in the lumen, which leads to limited zinc availability for this pathogen colonization in the gut [116-118].
On C difficile invasion, the intestinal immune system is activated, and calprotectin is released from the activated immune cells, particularly neutrophils, and eventually translocated across the intestinal epithelial cells. Calprotectin is a calcium- and zinc-binding protein, that can be used as an indirect marker of intestinal inflammation. Calprotectin released from the gut can bind zinc and limit the availability of zinc for pathogen colonization in the gut (Fig. 3) [109,116].
Another zinc transporter, ZIP8, is used as a host defense mechanism as well. A defect in ZIP8 gene impairs the host's defense against pneumonia because it can facilitate bacterial effects [119]. Variants of myeloid-specific ZIP8 can decrease the uptake of zinc and lead to intestinal microbiome dysbiosis and an increase in susceptibility to Streptococcus pneumoniae infection in the lung tissues of mice. In summary, zinc is a cofactor essential for microbiota-host communication. Possible perturbations in zinc homeostasis may impact the host immunity in the host and microbiome communities (Fig. 3) [120].
VitaminDWiki – Microbiome category contains
{include}
articles in both of the categories Microbiome and Gut
This list is automatically updated
{category}