The renaissance of vitamin D
Acta Biochim Pol. 2014 Dec 18. [Epub ahead of print]
Wierzbicka J1, Piotrowska A1, Żmijewski MA1.

Figure 1: Wire cage fixed to the tenement window, London, 1934.
There is no doubt that vitamin D plays a crucial role in the maintenance of musculoskeletal system. But the function of this ancient molecule presumably ranges far beyond hormone-like regulation, as it could be generated by simple unicellular organisms. First, we are going to discuss the role of vitamin D as a global regulator of homeostasis from a historical perspective, but later we will focus on current views and its relevance to human physiology and pathology. Three milestones are defining the impact of vitamin D on science and humanity.
Firstly, discovery that vitamin D is the cure for rickets, brought us supplementation programs and rapid irradiation of this devastating disease.
Secondly, detail description of photoproduction of vitamin D, its subsequent metabolism and interaction with vitamin D receptor VDR, provided mechanistic background for future discoveries.
Finally, recent large epidemiological studies provided indirect, but strong evidence that optimal level of vitamin D in serum has beneficial effects on our health and protects us from multiple diseases, including cancer.
Furthermore, existence of alternative pathways of vitamin D metabolism and multiple intracellular targets broadens our understanding of its physiological activities and offers new and very promising tools for prophylactics and treatment of many diseases of civilization.
Although vitamin D (and its derivatives) should not be regarded as a cure-all for every human disease, its beneficial effects on the human health have to be taken under consideration.
PMID: 25566549
📄 Download the uncorrected PDF from VitaminDWiki which follows
HISTORICAL PERSPECTIVE
Vitamin D is probably one of the oldest hormone that exists on earth (Bikle, 2011). Some of the earliest life forms have the capacity to produce vitamin D2, when exposed to the sunlight. Impressively, vitamin D2 producing phytoplankton survived, unchanged in the Atlantic Ocean, for more than 750 million years. There are many speculations, why these organisms need such excessive amounts of vitamin D2. One of the most probable is that ergosterol may acted as natural “sunscreen” protecting cells against an ultraviolet (UVA and UVB) radiation (Holick, 1989; Holick, 2003).
The modern history of vitamin D in human health began during Industrial Revolution, in the 17th century. City lifestyle and growing air pollution decreased an access to the sun resulting in massive outburst of rickets. In that time, the disease was recognized as a major health problem among children. Severe growth retardation, widening of the ends of the long bones, and bowing and bending of the legs are only a few clinical signs of rickets (Holick, 1994).
The importance of exposure to the sunlight and its association with the prevention and cure of this bone deforming disease in children was first recognized by Polish physician Jydrzej Sriadecki, in 1822. In his observations Sniadecki conchded that children living in the polluted, sunless centre of Warsaw (Poland), more often suffered from rickets than compared to children living in the sunny rural areas. He hypothesized, that lack of sufficient sun exposure was responsible for the development of rickets among children. What is more, he claimed that direct exposure to sunlight might be one of the most efficient methods to prevent and cure rickets. However, in the nineteenth century his studies were considered as incomprehensible and remained largely unnoticed. Of note, it was great Polish biochemist, Professor WVdz.mierz Mozolowski, who brought back Sniadecki's observations to the scientific community (Mozolowski, 1939).
Another scientist, who predicted the relationship between exposure to the sunlight and occurrence of rickets, was a British epidemiologist, Theobald Palm. In 1890, he combined notes from his travels with the opinions of colleagues and postulated that there is a negative dependence between latitude and occurrence of rickets. In his observations, he firmly highlighted that despite the poverty, in sunny, tropical areas there were significantly reduced number of rickets cases, than compared to “rich” urban countries. Similarly to Sniadecki, Palm was a supporter of the beneficial effects of the sun to bone health. He also strongly encouraged moving infants and children afflicted with rickets from large towns to sunnier rural areas (Palm, 1890). Once again, the observations and benefits arising from Palm’s findings remained unnoticed. It had taken another 30 years before significance of Sniadecki’s and Palm’s observations was explained.
In 1919, Huldschinsky showed that exposure to ultraviolet radiation from a mercury arc lamp resulted in regression of severe rickets. Furthermore, he demonstrated that the therapeutic effect of ultraviolet radiation was not limited to irradiate place, but had an equal effect on whole organism. For instance, the exposure to mercury arc lamp of just one arm resulted in the cure of rickets in both arms (Huldschinsky, 1919; Huldschinsky, 1928).
Two years after Huldschinsky findings, Hess and Unger conducted another valuable study. They exposed children suffering from rickets on a roof of a New York City hospital to sunshine for various periods. It resulted in significant improvement of the health of rachitic children as reported by X-ray examination (Hess & Unger, 1921). The results of above-mentioned studies prompted US government to establish an agency responsible for promoting an exposure of children to the sun in order to prevent rickets (Fig. 1). What is more, in the 1930s—1950s parents could buy ultraviolet lamps in local pharmacies to protect children from developing rickets (Hess, 1936; Eliot & Park, 1938). At that time, the popularity of UV irradiation was constantly growing. In 1924, Steenbock suggested that not only children and animals could be irradiated in order to prevent rickets but also irradiate food seems to be effective (Steenbock, 1924). This led to the UV irradiation of cows, their diet, and ultimately their milk to acquire food with antirachitic properties. Thanks to the fortification of milk with vitamin D rickets was very quickly eradicated in the United States and Europe. In the 1930s and 1940s in addition to milk, many other products fortified with vitamin D like bread, hot dogs, Twang soda, and even Schlitz beer were popular. Unfortunately, potential excessive consumption of vitamin D was blamed for the outbreak of hypercalcemia in Great Britain and US, in the 1940s. This resulted in tightening of the regulations concerning fortification of dairy products with vitamin D in Europe (Holick, 2006b).
Figure 1 . An example of the wire cage fixed to the tenement window, so that babies can benefit from sunshine. London, 1934.
Interestingly, it seems now that the outburst of hypercalcemia may not be attributed to an excessive supplementation, but also could be explained by coexistence of relatively rare diseases, including: Williams’ syndrome (Wacker & Holick, 2013), primary hyperparathyroidism (Michels & Kelly, 2013) or even sarcoidosis (Nunes et al., 2007). It could be also caused by specific mutation of CYP24A1 coding the main catabolic enzyme for vitamin D (Jacobs, 2014), resulting in abnormally high level of vitamin D in serum.
Simultaneously to ongoing studies on the effect of the sun on bone health, another potential antirachitic agent was found — a cod liver. The effectiveness of cod liver oil in prevention of the disease has been particularly valued on the coastlines of the Scandinavian countries and the United Kingdom (Ihde, 1975). Nevertheless, Schutte was a first physician, who prescribed cod liver oil as specific agent preventing rickets, in 1824 . From this time to the end of the century, German and French physicians have widely recommended cod liver oil as an antirachitic agent. Nevertheless, at the beginning of the 20th century, the usefulness of cod liver oil was questioned, probably due to a poor quality or impurity of prescribed cod liver oil (Guy, 1923). The first scientific approach to prove the anti-rachitic properties of cod liver oil was made by Edward Mellanby and Elmer McCollum. In classic, animal experiments, they accredited the antirachitic function of cod liver oil to fat-soluble vitamin A or other similar substance (Mellanby, 1919). However, in 1922, McCollum demonstrated that vitamin A do not possessed previously ascribed antirachitic functions. He was aware that oxidation destroys fat-soluble vitamin A, while similarly treated cod-liver oil preserved its protective action ag'inst the development of rickets. Thus, McCollum concluded that the antirachitic substance must be a distinct one from fat-soluble vitamin A. The newly discovered antirachitic factor from cod liver oil was named vitamin D, as it was fourth in the sequence of discovery of vitamins (McCollum et al., 1922).
In retrospect, the establishment of the fact that cod liver oil and sunlight were different but similar in their ability to prevent and treat rickets, was a significant advance in the study of vitamin D.
NATURALLY OCCURRING VITAMIN D ANALOGS
Although the idea of fat-soluble vitamin D as antirachitic factor became very clear at the beginning of the 20th century, the actual vitamin structure was not solved until 1932. First, Askew and co-workers succeeded in isolation of vitamin D2 (D2 or ergocalciferol) (Askew et al., 1931). Independently, a German group led by Win- daus in 1935 had isolated 7-dehydrocholesterol (Win- daus, Lettre & Schenck, 1935) and two years later vitamin D3 (Windaus & Bock, 1937). Interestingly, although Windaus was involved in discovery of vitamin D3 and its synthesis pathway, he received Nobel Prize in Chemistry on the account of his discoveries concerning structures of sterols and their relationship with vitamins, in 1928 (Wolf, 2004). Interestingly, he passed his patent's rights to the production of vitamin D by UV-irradiation of yeast derived ergosterol to Merck and Bayer companies. As a result, well known Vigantol is on the market since 1927 (Haas, 2007).
It was however still do not clear, whether the vitamin D is normally produced in the human body or could be found only in natural products. Henry Steen-bock was one of the pioneers in vitamin D research, who in an addition to early experiments with food irradiation (Steenbock, 1924) was later focused on the physiological activity of this fat-soluble vitamin. After his retirement in 1955, Hector DeLuca took over his laboratory and similarly to his predecessor, vitamin D became the centre of his attention. In 1968, he isolated an active vitamin D metabolite and identified it as 25-hydroxyvitamin D3 (25-OH D3) (Blunt et al, 1968). A few years later, he demonstrated that previously identified substance was produced in the liver (Gray et at., 1971). Further collaboration of Hector DeLuca with Michael Holick resulted in numerous discoveries. They identified the major circulating form of vitamin D, 25-hydroxyvitamin D3, (Holick et al, 1972a). Another achievement of a millstone was the discovery of biologically active metabolite of vitamin D — 1a,25-dihydroxyvitamin D3 (Holick et al, 1971). This was followed by the identification of other vitamin D metabolites, including: 24,25-dihydroxyvitamin D3, (Holick et al, 1972b), 1a,24,25-trihydroxyvitamin D3 (Holick et al, 1973) and 25,26-dihydroxyvitamin D3 (DeLuca et al, 1970). Those key discoveries enable us to understand the mechanisms associated with production and metabolism of vitamin D.
THE SKIN — FINAL LINK BETWEEN SUN AND VITAMIN D
Since Sniadecki’s times it was speculated that sun is essential for the product of the antirachitic factor, which was later described as vitamin D. However, it was Michael Holick, who for the first time showed effective synthesis of vitamin D in the skin subjected to ultraviolet radiation (Holick et al, 1977; Holick et al, 1979). His subsequent studies also confirmed experimentally, that the latitude or seasonal changes fect production of vitamin D in the skin (Webb et 1988). Interestingly, the influence of skin pigmentation on the efficiency of vitamin D production is still under debate . In individuals with black skin phenotype, the production of previtamin D3 was found Le be reduced to 20% of the white skin phototype (Fitzpatrick, 1988). More recent studies on the Us population showed that mean serum concentration; of 25(OH)D3 were approximately 25 nmol/L less in African-Americans than in Caucasian (Looker et m., 2008). Therefore, darker-skinned individuals require longer exposures to achieve the same plasm a 25(OH)D3 concentration. However, even people with highly pigmented skin can obtain relatively high 25(OH)D3 concentrations in the serum, as it was observed among Gambia population (Prentice, 2008).
Interestingly, it was the production in the skin subjected to the sunlight not supplementation, that was found to be the most efficient source of vitamin D. The exposure of the skin to only 1 minimal erythemal dose (MED) of the sunlight results in production of at least 20 000 Units of vitamin D (Holick, 2008). Furthermore, skin production of vitamin D does not cause the symptoms of an overdose, because the excessive exposition to UVB light leads to its photodegradation. The main products are 5,6-transvitamin D3, and suprasterols I and II (Webb et al, 1989). Other photoproducts including 5,7,9(11)-trienes were described, recently (Chignell et al, 2006; Zmijewski et al, 2009).
Thus, almost 250 years after Sniadecki's observations, it became obvious that beneficial role of the sunlight in rickets is attributed to skin production of vitamin D3.
CLASSIC PATHWAY OF PRODUCTION AND METABOLISM OF VITAMIN D
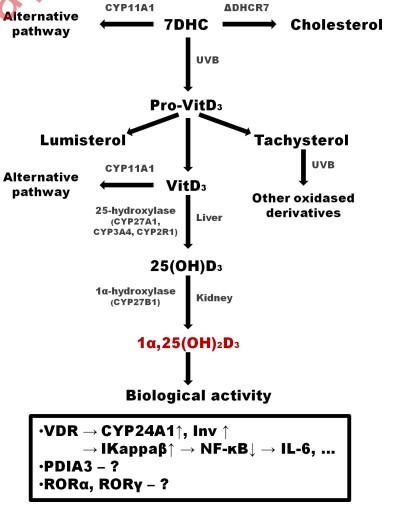
It is well established that a biologically active form of vitamin D3 is 1a,25(OH)2D3 (Fig. 2). It is produced in multistep process involving photochemical isomerisation of 7-dehydrocholesterol (cholesta-5 ,7-dien-3 -ol, 7-DHC) followed by enzymatic hydroxylation of vitamin D. Initially, under the UVB radiation, the B-ring of 7-DHC is photolysed, what leads to formation of previtamin D3 (Holick et al, 1977). The reaction takes place in keratinocytes of the base layer of the epidermis (Holick et al, 1977; Slominski et al, 2012c). Then, previtamin D3 isomerizes to vitamin D3, tachysterol3 (T3) and lumister- ol3 (L3). Vitamin D3 can be released to the circulation, where it is transported by vitamin D-binding protein (DBP) (Lehmann, 2009). Circulating vitamin D is activated by subsequent hydroxylations. First hydroxylation to 25-hydroxycholecalciferol (25-OH D3) takes place in the liver and is carried out by mitochondrial or microsomal 25-hydroxylases (CYP2j!1, CYP27A1). Next, in the kidneys 25-OH D3 i s hydroxylated by mitochondrial 1 a-hydroxylase (CYP27B1) to 1a,25(OH)2D3 (calcitriol) — the fully active form of vitamin D3 (Takeyama et al, 1997). Addnonally, a number of distinct tissues and organs, such as intestines, also have the ability to activate vitamin D by its hydroxylation (Hewison et al, 2004). It should be emphasized, that the skin is the only known organ equipped with complete machinery of vitamin D3 production and metabolism including enzymes responsible for 25- and 1a-hydroxylation as well as the vitamin D receptor (Bouillon et al, 2008; Luderer & De- 2010). The level of 25-OH D3 and 1a,25(OH)2D3 in circulation and tissues is regulated by 24-hydroxylase (CYP24A1), which transforms them to 24,25(OH)2D3 or 1a,24,25(OH)3D3, respectively. Further catabolism results in formation of water-soluble calcitroic acid, which is exerted in urine (Reddy & Tserng, 1989).
Vitamin D2 is exclusively formed from ergosterol by fungi and phytoplankton subjected to UVB radiation (Holick et al., 1982). Some plants have also limited capacity to produce vitamin D2 (Japelt et aL, 2013). Similarly to 7-DHC, the photolysis of ergosterol results in formation of three main products: vitamin D2, lumisterol2 and tachysterol2 (Kalaras et al, 2012). Furthermore, acquired vitamin D2 undergoes hydroxylation at position C25 in the liver and C1 in the kidney to produce biologically active 1,25(OH)2D2 (Holick, 2003; Zhu & DeLuca, 2012). It has to be added, that vitamin D2 is a major form of dietary vitamin D in humans, especially in Western Europe and USA (Holick, 2003; Bikle, 2011). However, the question whether vitamin D2 is an ideal replacement for D3 is still open to debate (Leventis & Kiely, 2009).
ALTERNATIVE PATHWAY LEADING TO NOVEL VITAMIN D ANALOGS
Although classical pathway of vitamin D synthesis and metabolism was established long time ago, new class of vitamin D derivatives was recently described. Collaboration of Andrzej Slominski and Robert Tuckey (recent reviewed Slominski et al, 2013b) resulted in discovery of novel metabolic pathway for 7-DHC and vitamin D in animals (Fig. 2). The alternative route is initiated by the enzymatic action of cytochrome P450scc (CYP11A1) on 7-dehydrocholesterol (Slominski et al, 2004; Slominski, Kim et al., 2013a; Slominski et al., 2014). It was shown conversion of cholesterol to pregnenolone may catalyse the transformation of 7-DHC to 7-DHP hydroprognenolone). This conversion requires hydroxy lation of 7-DHC at the C22 and C20 positions, followed by cleavage of side chains resulting in formation of 7-DHP (Slominski et al., 2004). It seems that /-DHP may be further modified by classical steroid metabolic enzymes (17a-hydroxylase and 17, 20-liase), leading to the formation of new steroidal 5,7-dienes with modified side chains (Slominski et al., 2009). All of those compounds can serve as precursors for the vitamin D-like derivatives after exposure to the UVB radiation (Zmi- jewski et al., 2008; Zmijewski et al., 2009; Zmijewski et al., 2011). Interestingly, not only 7-DHC but also ergos- terol, vitamin D3 and vitamin D2 can be metabolise by cytochrome P450scc,what results in formation of a new class of hydroxylderivatives (Slominski et al., 2005b; Slominski et al., 2005a; Slominski et al, 2006; Slominski et al, 2013a).
The CYP11A1 was shown also to act on C20 of vitamin D analogs (e.g. vitamin D2 and D3) generating family of novel vitamin D hydroxyderivatives (Slominski et al., 2005; Tuckey et al., 2008). Furthermore, it was shown that major product of hydroxylation of vitamin D, namely 20-hydroxyvitamin D3(20-OH D3) (Guryev et al., 2003; Slominski et al., 2005b) can be hydroxylated by CYP11A1 to 20,23-dihydroxyvitamin D3 (20,23(OH)2D3) (Slominski et al, 2005b; Tuckey et al, 2011). It has to be stress out that CYP11A1-initiated metabolism of vitamin D3 was detected both, in vitro (Tuckey et al., 2008) and in vivo (Slominski et al., 2012a). Futhermore, several new studies showed that Cyp450scc generated vitamin D hydroxyderivatives are biologically active. Importantly, it was shown that they are less prone to induce hypercalcemia, therefore they are currently investigated as potential anti-leukemia (Slominski et al., 2010) and anti-melanoma factors (Slominski et al., 2012a). Moreover, 20-OH D3 exhibits anti-proliferative and pro-differentiation activities in human epidermal keratinocytes, as it was shown recently (Zbytek et al., 2008; Slominski et al, 2011). Nevertheless, further research is needed in order to establish physiological role of alternative vitamin D metabolites and their potential applications in therapy (recent review: Szyszka et al, 2012).
INTERCELLULAR MECHANISM ACTIVATED BY VITAMIN D AND ITS ANALOGS
Final puzzle in the studies on vitamin D was to establish, how it is possible that one, relatively simple molecule expresses such a variety of biological functions. It turns out that similarly to other steroid hormones, vitamin D activates its canonical nuclear receptor. Exploration of the classical pathway began in 1969, when Haussler and Norman discovered the nuclear receptor for 1,25(OH)2D3 (Haussler & Norman, 1969). Over the following years, scientists revealed expression of the vitamin D receptor (VDR) in many target tissues (Cavalier, 2009). More than twenty years later VDR expression was also shown in epidermal keratinocytes (Milde et al., 1991). According to well-established “genomic pathway”, vitamin D exerts its biological activity by binding with nuclear receptor — VDR, which after stimulation forms a dimer with 9-cis-retinoic acid receptor — RXR (retinoid X receptor). The complex is then translocated to nucleus and acts as a transcriptional factor by bind- sors interact with VDR-RXR complex and regulates its activity (Silvagno et al., 2013). Initially, VDRE was discovered in the promoter region of bone-specific osteocalcin gene (Morrison et al, 1989). It seems that this was symptomatic for the vitamin D history. Since then, the VDR action as transcription factor was extensively investigated by both mRNA and miRNA microarrays. For example, in squamous cell carcinoma cells over 900 genes was shown to respond to 1a,25(OH)2D3 (Wang et al., 2005). It was demonstrated that 1a,25(OH)2D3 can regulate genes controlling extracellular matrix structure and its remodelling, cell adhesion or inducting a basal keratinocyte phenotype (Lin et al., 2002).
Having in mind ancient origin of vitamin D, its interaction with VDR seems to be quite recent adapted pathway regulating its activity. In fact, so-called, nongenomic mechanism of rapid vitamin D response has been described recently. This mechanism does not directly affect gene expression or require additional protein synthesis. Rapid vitamin D response was shown to modulate intracellular calcium levels, affects activity of several intracellular signalling pathways, through activation of selected phosphate kinases and phosphatases. These activities take minutes and occur in the cytoplasm of the cell rather than in the nucleus. Potential mechanism of non-genomic response involves interaction of vitamin D to 1a,25(OH)2D membrane—associated rapid response steroid-binding protein (1,25 D-MARRSBP), also known as the protein-disulfideisomerase-associated 3 (PDIA3) or endoplasmic reticulum stress protein 57 (ERp57) (Ne- mere et al, 2004). PDIA3 activates phospholipase C in a G protein-coupled process and results in production of inositol trisphosphate (IP3) and diacylglycerol. These two cellular messengers mediate the rapid release of calcium from the cellular stores (Nemere et al, 2012).
Recently, new targets for vitamin D have been discovered. It was that secosteroids (such as 20-OH D3 and 20,23(OH)2D3) can act as antagonists of retinoic acid-related orphan receptors a and y (RORa and RORy) (Slominski et al., 2014). RORa and RORy are the members of the nuclear receptors ROR subfamily, which take part in the regulation of a number of physiological processes — affects several immune functions, metabolism, and cerebellar development (Jetten, 2009). RORs are expressed in a variety of tissues, including testis and kidneys (Jetten, 2009). Moreover, their presence was also confirmed in human skin cells (Slominski et al, 2014). Crystallography provided insightful structure of the ligand binding pockets of RORs (Stehlin et al., 2001). Those observations not only confirmed that RORs can function as the ligand-dependent transcription factors, but also imply that RORs might be new interesting therapeutic targets for vitamin D analogs. Especially if we take under consideration several reports indicating a potential role of RORs in osteoporosis, autoimmune diseases, asthma, cancer, and obesity (Jetten, 2009). Thus, it seems that our relatively simple model for vitamin D, shown on Fig. 2 is going to be modified in forthcoming years in order to explain complexity of vitamin D metabolism and its pleiotropic activities.
THE RENAISSANCE OF VITAMIN D3
The classic physiologic function of vitamin D3 is to maintain calcium and phosphorus homeostasis, ensuring proper metabolic functions of bone mineralization and neuromuscular transmission (Holick, 2006a). Until recently, vitamin D was considered as “the bone vitamin diseases such as rickets or osteomalacia. As a result vitamin D discovery, rickets was successfully eradicated by proper supplementation (Holick, 2013). However, it has to be underline that vitamin D is also crucial for maintenance of nervous and cardiovascular and immunological systems, as well as plays important role in skin physiology, to name only a few. Recent 20 years brought us constantly increasing number of epidemiological studies indicating that maintenance of the optimal level of 25-OH D3 in the serum is simply essential for our health. Amongst others, vitamin D was found to be a protecting agent against multiple types of cancer (Garland et al, 2006), bacterial infections (Bikle, 2008); autoimmune (Munger et al., 2006) or cardiovascular diseases (Wang et al., 2008; Tukaj et al., 2012). It seems that vitamin D affects at least in part all major human function at the cellular, organ and whole body levels (Fig. 3). Multiple epidemiological studies provide strong evidence that monitoring of vitamin D status is the key factor in proper supplementation. For instance, recent Polish study (Gdansk region) showed that amongst 448 volunteers only 2.5% had optimal concentration of 25- OH D3 in serum (more thar 30 ng/gL) in winter of 2012. Moreover, lack of proper supplementation and food fortification with vitamin D in Poland resulted in very low mean of 25-OH D3 concentration (14.3 ± 6.6 ng/gL). Interestingly, only individuals with recent episodes of UVB exposure had sufficient concentration of vitamin D (Kmiec, 2014). This finding as many other, strongly suggested the necessity of proper supplementation (Webb et , 2010; Trofimiuk-Muldner et al., 2012). Furthermore, routinely suggested 400—800 Units of vitamin D, pe" day, may not be sufficient for efficient supplementation. Thus, higher doses up to 2000 U are recommended according to recently published guidance for Central Europe (Pludowski, 2013). This detail recommendation concerning supplementation for different goups was published, recently in English (Pludowski t a’., 2013a) and in Polish as well (Pludowski et al., 13b). It has to be also mentioned that monitoring of optimal supplementation. It is especially important for several groups of individuals vulnerable to vitamin D deficiency, including pregnant woman, children, and el-
CONCLUSIONS
In spite of more than 100 years published articles, the overall impact its derivatives on human health is not fully understood. Having in mind its ancient origin, it is obvious that vitamin D regulates multiple pathways and has an impact on human physiology and internal homeostasis. Proper activity of multiple organs depends on optimal vitamin D level, thus it should be not a surprise that vitamin D deficiency is an important factor involved in development of multiple human diseases.
Recent studies revealing existence of alternative pathways of vitamin D metabolism and activity broaden our knowledge concerning pleiotropic impact of vitamin D and its derivatives on human physiology and pathology. Furthermore, newly discovered low-calcemic vitamin D analogs such as 20-OH D3; or intracellular targets like PDIA3 or RORs, provide new opportunities for the therapy of multiple human diseases.
Acknowledgements
Supported by grant of Polish Ministry of Science and Higher Education, project no. N405 623238 and N402 662840 (MAZ).
References
Askew FA, Bourdillon RB, Bruce HM, Jenkins RGC, Webster TA (1931) The distillation of vitamin D. Proc Biol Sci 107: 76-90.
Bikle DD (2008) Vitamin D and the m une system: role in protection against bacterial infection. Cur OPin Nephrol Hypertens 17: 348-352.
Bikle DD (2011) Vitamin D: an an .rent hormone. Exp Dermatol 20: 7-13.
Blunt JW, DeLuca HF, ch'ops HK (1968) 25-hydroxycholecalciferol. A biologically active metabolite of vitamin D3. Biochemistry 7: 3317-3322.
Bouillon R, Carmeliet G, Verlinden L, van Etten E, Verstuyf A, Lu- derer HF, Lieben L, Mathieu C, Demay M (2008) Vitamin D and human health: lessons from vitamin D receptor null mice. Endocr Rev 29: 726-76.
Cavalier E, Dela naye P, Chapelle JP, Souberbielle JC (2009) Vitamin D: current status and perspectives. Clin Chem Lab Med 47: 120-127.
Chignell CF, Kukielczak BM, Sik RH, Bilski PJ, He YY (2006) Ul- travmlet A sensitivity in Smith-Lemli-Opitz syndrome: Possible in- ement of cholesta-5,7,9(11)-trien-3 beta-ol. Free Radic Biol Med 39-346.
Chez. . walska J, Pludowski P, Dobrzanska A, Kryskiewicz E, Karc- arewicz E, Gruszfeld D, Pleskaczynska A, Golkowska M (2012) pact of vitamin D supplementation on markers of bone mineral tabolism in term infants. Bone 51: 781-786.
Deluca HF, Suda T, Schnoes HK, Tanaka Y, Holick MF (1970) 25,26-dihydroxycholecalciferol, a metabolite of vitamin D3 with intestinal calcium transport activity. Biochemistry 9: 4776-4780.
Eliot MM, Park EA (1938) Brennemann’s Practice of Pediatrics. In Rickets, vol 1, pp 1-110. W.F. Prior Company, Inc.
Feldman D, Krishnan AV, Swami S, Giovannucci E, Feldman BJ (2014) The role of vitamin D in reducing cancer risk and progression. Nat Rev Cancer 14: 342-357.
Fitzpatrick TB (1988) The validity and practicality of sun-reactive skin types I to VI. Arch Dermato 124: 869-871.
Garland CF, Garland FC, Gorham ED, Lipkin M, Newmark H, Mohr SB, Holick MF (2006) The role of vitamin D in cancer prevention. Am J Public Health 96: 252-261.
Gray R, Boyle I, DeLuca HF (1971) Vitamin D metabolism: the role of kidney tissue. Science 172: 1232-1234.
Guy RA (1923) The history of cod liver oil as a remedy. Am J Dis Child 26: 112-116.
Haas J (2007) Vigantol-Adolf Windaus and the history of vitamin D. Wurzbg Medizinhist Mitt 26: 144-181.
Hansen CM, Hamberg KJ, Binderup E, Binderup L (2000) Seocalci- tol (EB 1089): a vitamin D analogue of anti-cancer potential. Background, design, synthesis, pre-clinical and clinical evaluation. Curr Pharm Des 6: 803-828.
Haussler MR, Norman AW (1969) Chromosomal receptor for a vitamin D metabolite. Proc Natl Acad Sci 62: 155-162.
Hess AF, Unger LJ (1921) The cure of infantile rickets by sunlight. J Am Med Ass 77: 39-41.
Hess AF (1936) In Collected Writings, volume 1. Thomas CC, eds, pp 669-719. Springfield, IL.
Hewison M, Zehnder D, Chakraverty R, Adams JS (2004) Vitamin D and barrier function: a novel role for extra-renal 1 alpha-hydroxylase. Mol Cell Endocrinol 215: 31-38.
Holick MF, Schnoes HK, DeLuca HF, Suda T, Cousins RJ (1971) Isolation and identification of 1,25-dihydroxycholecalciferol. A metabolite of vitamin D active in intestine. Biochemistry 10: 2799-2804.
Holick MF, DeLuca HF, Avioli LV (1972a) Isolation and identification of 25-hydroxycholecalciferol from human plasma. Arch Intern Med 129: 56-61.
Holick MF, Schnoes HK, DeLuca HF, Gray RW, Boyle IT, Suda T (1972b) Isolation and identification of 24,25-dihydroxycholecalcif- erol, a metabolite of vitamin D made in the kidney. Biochemistry 11: 4251-4255.
Holick MF, Kleiner-Bossaller A, Schnoes HK, Kasten PM, Boyle IT, DeLuca HF (1973) 1,24,25-Trihydroxyvitamin D3. A metabolite of vitamin D3 effective on intestine. J Biol Chem 248: 6691—6696.
Holick MF, Frommer JE, McNeill SC, Richtand NM, Henley JW, Potts JT Jr (1977) Photometabolism of 7-dehydrocholesterol to previtamin D3 in skin. Biochem Biophys Res Commun 76: 107—114.
Holick MF, Richtand NM, McNeill SC, Holick SA, Frommer JE, Henley JW, Potts JT Jr. (1979) Isolation and identification of previtamin D3 from the skin of rats exposed to ultraviolet irradiation. Biochemistry 18: 1003—1008.
Holick MF, Holick SA, Guillard RL (1982) Photosynthesis of previtamin D in phytoplankton. In Current Trends in Comparative Endocrinology. Lofts B, Holmes WN, eds. Hong Kong University Press, Hong Kong.
Holick MF (1989) Phylogenetic and evolutionary aspects of vitamin D from phytoplankton to humans. In Vertebrate Endocrinology: Fundamentals and Biomedical Implications. Pang PKT ed, vol 3, pp 7—43. Academic Press, Orlando, FL.
Holick MF (1994) McCollum Award Lecture, 1994: vitamin D new horizons for the 21st century. Am J Clin Nutr 60: 619—630.
Holick MF (2003) Vitamin D: A millenium perspective. J Cell Biochem 88: 296-307.
Holick MF (2004) Vitamin D: importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am J Clin Nutr 79: 362-371.
Holick MF (2006 a) High prevalence of vitamin D inadequacy and implications for health. Mayo Clin Proc 81: 353-373.
Holick MF (2006b) Resurrection of vitamin D deficiency and rickets. J Clin Invest 116: 2062-2072.
Holick MF (2008) Sunlight, UV-radiation, vitamin D and skin cancer: how much sunlight do we need? Adv Exp Med Biol 624: 1-15.
Holick MF (2013) Vitamin D: A milenium perspective. J Cell Biochem 88: 296-307.
Huldschinsky K (1919) Heilung von Rachitis durch Kunstliche Hohen- sonne (Ultraviolet irradiation of rachitic children). Deutsche Medizinis- che Wochenschrift 45: 712-713.
Huldschinsky K (1928) The Ultra-violet light treatment of rickets. Alpine Press 3-19.
Ihde AJ (1975) Studies on the history of rickets. II. The roles of cod liver oil and light. Pharm Hist 17: 13-20.
Jacobs TP, Kaufman M, Jones G, Kumar R, Schlingmann KP, Shapses S, Bilezikian JP (2014) A lifetime of hypercalcemia and hypercalciu- ria, finally explained. J Clin Endocrinol Metab 99: 708-712.
Japelt RB, Jakobsen J (2013) Vitamin D in plants: a review of occurrence, analysis, and biosynthesis. Front Plant Sci 13: 136.
Jetten AM (2009) Retinoid-related orphan receptors (RORs): roles in development, immunity, circadian rhythm, and cellular tabolism. Nucl Recept Signal 7: e003.
Kalaras MD, Beelman RB, Holick MF, Elias RJ (2012) Genei potentially bioactive ergosterol-derived products follown ultraviolet light exposure of mushrooms (Agaricus bispor Chem 135: 396-401.
Kamel K, Kolinski A (2012) Assessment of the free billing energy of 1,25-dihydroxyvitamin D3 and its analogs with a.' human VDR receptor model. Acta Biochim Pol 59: 653-660.
Karczmarewicz E, Czekuc-Kryskiewicz E, PludowAi P (2013) Effect of vitamin D status on pharmacological treatment efficiency. Derma- toendocrinol 5: 299-304.
Kmiec P, Zmijewski MA, Waszak P, Swormak K, Lizakowska-Kmiec M (2014) Vitamin D deficiency during- winter months among an adult, predominantly urban, population in Northern Poland. En- dokrynol Pol 65: 105-113.
Lehmann B (2009) Role of the vitamin D3 pathway in healthy and diseased skin — facts, contradictions and hypotheses. Exp Dermatol
Mellanby E (1919) An experimental investigation on rickets. Lancet 1: 407-412.
Milczarek M, Chodynski M, Filip-Psurska B, Martowicz A, Krupa M, Krajewski K, Kutner A, Wietrzyk J (2013) Synthesis and biological activity of diastereomeric and geometric analogs of calcipotriol, PRI-2202 and PRI-2205, against human HL-60 leukemia and MCF- 7 breast cancer cells. Cancers (Basel) 5: 1355-1378.
Milczarek M, Filip-Psurska B, Swietnicki W, Kutner A, Wietrzyk J (2014) Vitamin D analogs combined with 5-fluorouracil in human HT-29 colon cancer treatment. Oncol Rep 32: 491-504.
Milde P, Hauser U, Simon T, Mall G, Ernst V, Haussler MR, Frosch P, Rauterberg EW (1991) Expression of 1,25-dihydroxyvitamin D3 receptors in normal and psoriatic skin. J Invest Dermatol 97: 230-239.
Morrison NA, Shine J, Fragonas JC, Verkest V, McMenemy ML, Ei- sman JA (1989) 1,25-dihydroxyvitamin D-responsive element and glucocorticoid repression in the osteocalcin gene. Science 246: 11581161.
Mozolowski W (1939) Sniadecki J. Jerdrzej Sniadecki (1768—1838) on the cure of rickets. Nature 14: 121-124.
Munger KL, Levin LI, Hollis BW, Howard NS, Ascherio A (2006) Serum 25-hydroxyvitamin D levels and risk of multiple sclerosis. J Am Med Ass 296: 2832-2838.
Nemere I, Farach-Carson MC, Rohe B, Sterling TM, Norman AW, Boyan BD, Safford SE (2004) Rmo'yme knockdown functionally links a 1,25(OH)2D3 membrane bind’ng protein (1,25D3-MARRS) and phosphate uptake in intestinal cells. Proc Natl Acad Sci USA 101: 7392-7397.
Nemere I, Garbi N, Hammering G, Hintze KJ (2012) Role of the 25D3-MARRS receptor in the 1,25(OH)2D3-stimulated uptake of calcium and phosphate -n intestinal cells. Steroids 77: 897-902.
Nunes H, Bouvry D, S'Vr P, Valeyre D (2007) Sarcoidosis. Orphanet J Rare Dis 2: 46.
Palm T (1890) The geographical distribution and etiology of rickets. Practitioner 45: 270-279, 321-342.
Pludowski P, Karczmarewicz E, Bayer M, Carter G, Chlebna-Sokol D, Czech-Kowalska J, Debski R, Decsi T, Dobrzanska A, Franek E, Gluszko P, Grant WB, Holick MF, Yankovskaya L, Konstantynow- icz J, Ksiazyk JB, Ksiezopolska-Orlowska K, Lewinski A, Litwin M, Lohner S, Lorenc RS, Lukaszkiewicz J, Marcinowska-Suchowierska wicz A, Misiorowski W, Nowicki M, Pilz S, Povoroznyuk V, t P, Rudenka E, Shoenfeld Y, Socha P, Solnica B, Szalecki 'alalaj M, Varbiro S, Zmijewski M (2013a) Practical guidelines :he supplementation of vitamin D and the treatment of deficits in Central Europe — recommended vitamin D intakes in the general population and groups at risk of vitamin D deficiency. Endokrynol Pol 64: 319-327.
Pludowski P, Karczmarewicz E, Chlebna-Sokol D, Czech-Kowalska J, Debski R, Dobrzanska A, Franek E, Gluszko P, Konstantynowicz
J, Ksiazyk JB, Ksiezopolska-Orlowska K, Lewinski A, Litwin M, Lorenc RS, Lukaszkiewicz J, Marcinowska-Suchowierska E, Mile- wicz A, Misiorowski W, Nowicki M, Rozentryt P, Socha P, Sol- nica B, Szalecki M, Talalaj M, Zmijewski MA (2013b) Witamina D: rekomendacje dawkowania w populacji osob zdrowych oraz w grupach ryzyka deficytow: wytyczne dla Europy Srodkowej 2013 r. Standardly Med Pediatria 10: 573-578 (in Polish).
Prentice A (2008) Vitamin D deficiency: a global perspective. Nutr Rev 66: S153-S164.
Reddy GS, Tserng KY (1989) Calcitroic acid, end product of renal metabolism of 1,25-dihydroxyvitamin D3 through C-24 oxidation pathway. Biochemistry 28: 1763-1769.
Silvagno F, Consiglio M, Foglizzo V, Destefanis M, Pescarmona G (2013) Mitochondrial translocation of vitamin D receptor is mediated by the permeability transition pore in human keratinocyte cell line. PLoS One 8: e54716.
Slominski A, Zjawiony J, Wortsman J, Semak I, Stewart J, Pisarchik A, Sweatman T, Marcos J, Dunbar C, Tuckey RC (2004) A novel pathway for sequential transformation of 7-dehydrocholesterol and expression of the P450scc system in mammalian skin. Eur J Biochem 271: 4178-4188.
Slominski A, Semak I, Zjawiony J, Wortsman J, Gandy MN, Li J, Zbytek B, Li W, Tuckey RC (2005a) Enzymatic metabolism of ergo sterol by cytochrome p450scc to biologically active 17alpha,24- dihydroxyergosterol. Chem Biol 12: 931-939.
Slominski A, Semak I, Zjawiony J, Wortsman J, Li W, Szczesniewski A, Tuckey RC (2005b) The cytochrome P450scc system opens an alternate pathway of vitamin D3 metabolism. FEBS J 272: 40804090.
Slominski A, Semak I, Wortsman J, Zjawiony J, Li W, Zbytek B, Tuckey RC (2006) An alternative pathway of vitamin D metabolism. Cytochrome P450scc (CYP11A1)-mediated conversion to 20-hydroxy- vitamin D2 and 17,20-dihydroxyvitamin D2. FEBS J 273: 2891-901. Slominski AT, Zmijewski MA, Semak I, Sweatman T, Janjetovic Z, Li W, Zjawiony JK, Tuckey RC (2009) Sequential metabolism of 7-dehydrocholesterol to steroidal 5,7-dienes in adrenal glands and its biological implication in the skin. PLoS One 4: e4309.
Slominski AT, JanjetovicZ, Fuller BE, Zmijewski MA, Tuckey RC, Nguyen MN, Sweatman T, Li W, Zjawiony J, Miller D, Chen TC, Lozanski G, Holick MF (2010) Products of vitamin D3 or 7-dehydrocholesterol metabolism by cytochrome P450scc show antileukemia effects, having low or absent calcemic activity. PLoS One 5: e9907.
Slominski AT, Janjetovic Z, Kim TK, Wright AC, Grese LN, Riney SJ, Nguyen MN, Tuckey RC (2012a) Novel vitamin D hydroxyderiva- tives inhibit melanoma growth and show differential effects on normal melanocytes. Anticancer Res 32: 3733—3742.
Slominski A, Kim TK, Chen J, Nguyen MN, Li W, Yates CR, Sweat- man T, Janjetovic Z, Tuckey RC (2012b) Cytochrome P450scc-de- pendent metabolism of 7-dehydrocholesterol in placenta and epidermal keratinocytes. Int J Biochem Cell Biol 44: 2003—2018.
Slominski AT, Kim TK, Shehabi HZ, Semak I, Tang EK, Nguyen MN, Benson HA, Korik E, Janjetovic Z, Chen J, Yates CR, Postlethwaite A, Li W, Tuckey RC (2012c) In vivo evidence for a novel pathway of vitamin D(3) metabolism initiated by P450scc and modified by CYP27B1. Faseb J 26: 3901-3915.
Slominski A, Kim TK, Li W, Yi AK, Postlethwaite A, Tuckey RC (2013a) The role of CYP11A1 in the production of vitamin D metabolites and their role in the regulation of epidermal functions. J Steroid Biochem Mol Biol 144: 28-39.
Slominski A, Kim TK, Zmijewski MA, Janjetovic Z, Li W, Chen J, Kusniatsova EI, Semak I, Postlethwaite A, Miller DD, Zjawiony JK, Tuckey RC (2013b) Novel vitamin D photoproducts and their precursors in the skin. Dermatoendocrinol 5: 7-19.
Slominski AT, Kim TK, Takeda Y, Janjetovic Z, Brozyna AA, Skob- owiat C, Wang J, Postlethwaite A, Li W, Tuckey Rc, Jetten AM (2014) RORalpha and ROR gamma are expressed in human skin and serve as receptors for endogenously produced noncalcemic 20-hydroxy- and 20,23-dihydroxyvitamin D. Faseb J 28: 2775-2789.
Steenbock H (1924) The induction of growth promoting and calcifying properties in a ratio by exposure to light. Science 60: 224-225.
Stehlin C, Wurtz JM, Steinmetz A, Greiner E, Schule R, Moras D, Renaud JP (2001) X-ray structure of the orphan nuclear receptor RORbeta ligand-binding domain in the active conformation. Embo J 20: 5822-5831.
Szyszka P, Zmijewski MA, Slominski AT (2012) New vitamin D analogs as potential therapeutics in melanoma. Expert Rev Anticancer Ther 12: 585-599.
Takeyama K, Kitanaka S, Sato T, Kobori M, Yanagisawa J, Kato S (1997) 25-Hydroxyvitamin D3 1 alp ha-hydroxylase and vitamin D synthesis. Science 211: 1827-1830.
Trofimiuk-Muldner M, Kiec-Klimczak M, Hubalewska-Dydejczy (2012) Niedobor witaminy D w populacji miasta Krakowa - ki wst^pne. Endokrynol Pol 63: 160 (in Polish).
Tuckey RC, Li W, Shehabi HZ, Janjetovic Z, Nguyen MN, Kim TK, Chen J, Howell DE, Benson HA, Sweatman T, Baldisseri DM, Slo- minski A (2011) Production of 22-hydroxy metabolites of vitamin d3 by cytochrome p450scc (CYP11A1) and analysis of their biological activities on skin cells. Drug Metab Dispos 39: 1577-1588.
Tukaj S, Trzonkowski P, Tukaj C (2012) Regulatory effects of 1,25-di- hydroxyvitamin D3 on vascular smooth muscle cells. Acta Biochim Pol 59: 395-400.
Wacker M, Holick MF (2013) Sunlight and Vitamin D: A global perspective for health. Dermatoendocrinol 5: 51-108.
Wang TJ, Pencina MJ, Booth SL, Jacques PF, Ingelsson E, Lanier K, Benjamin EJ, D'Agostino RB, Wolf M, Vasan RS (2008) Vitamin D deficiency and risk of cardiovascular disease. Circulation 111: 503511.
Wang TT, Tavera-Mendoza LE, Laperriere D, Libby E, MacLeod NB, Nagai Y, Bourdeau V, Konstorum A, Lallemant B, Zhang R, Mader S, White JH (2005) Large-scale in silico and microarray-based identification of direct 1,25-dihydroxyvitamin D3 target genes. Mol Endocrinol 19: 2685-2695.
Webb AR, Kline L, Holick MF (1988) Influence of season and latitude on the cutaneous synthesis of vitamin D3: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D3 synthesis in human skin. J Clin Endocrinol Metab 61: 373-378.
Webb AR, DeCosta BR, Holick MF (1989) Sunlight regulates the cutaneous production of vitamin D3 by causing its photodegradation. J Clin Endocrinol Metab 68: 882-887.
Webb AR, Kift R, Durkin Mi. O'Brien SJ, Vail A, Berry JL, Rhodes LE (2010) The role of sunlight exposure in determining the vitamin D status of the UK white adult population. Br J Dermatol 163: 1050-1055.
Windaus A, Lettre H, Schenck F (1935) 7-dehydrocholesterol. Ann Chem 520: 98-107.
Windaus A, Bock F (1937) U" ber das provitamin aus dem sterin der schweineschwarte. Z Physiol Chem 245: 168-170.
Wolf G (2004) ”he discovery of vitamin D: the contribution of Adolf Windaus. j Nutr 134: 1299-302.
Zhu J, DeLuca HF (2012) Vitamin D 25-hydroxylase — four decades of se. rching, are we there yet? Arch Biochem Biophys 523: 30-36.
Zmijewsk. MA, Li W, Zjawiony JK, Sweatman TW, Chen J, Miller lominski AT (2008) Synthesis and photo-conversion of an- and pregna-5,7-dienes to vitamin D 3-like derivatives. Photo-m Photobiol Sci 1: 1570-1576.
Zmijewsk MA, Li W, Zjawiony JK, Sweatman TW, Chen J, Miller Slominski AT (2009) Photo-conversion of two epimers (20 R and 20S) of pregna-5,7-diene-3beta, 17alpha, 20-triol and their bioactivity in melanoma cells. Steroids 14: 218-228.
See also VitaminDWiki
Vitamin D history back to Egyptians and fortification - Aug 2011
More sunshine: more teeth (1862), fewer cavities (1934) – Oct 2011
History of vitamin D – Nov 2012 Dr. Norman
History of rickets and sunshine in a bottle then vitamin D – Jan 2012
SOLAR ultraviolet radiation and vitamin D: a historical perspective – Aug 2007
Vitamin D and its active metabolites – DeLuca Jan 2014 History