Many Vitamin D similarities: people and cows
A few of the many Vitamin D similarities in the study
If less time in the sun, more vitamin D supplementation (or UVB) is needed
The amount of Vitamin D needed varies with the individual
Need a minimum of 30 IU /kilogram of body weight
which would be 10,000 IU for a 150 lb human
30 ng is minimum acceptable, 70 ng is much better
200 ng is not a problem
A good level of vitamin D is vital for the immune system
Cofactors are important
Vitamin D Receptor is important ( VDR )
The same genes can limit vitamin D in the blood (such as DBP )
Vitamin D is stored in the muscle
Please email [hlahore@gmail.com] with sentences in the study which have additional simularities
Other readers would like to know about them and I only had time to skim this study
Vitamin D in dairy cows: metabolism, status and functions in the immune system
Archives of Animal Nutrition, DOI: 10.1080/1745039X.2021.2017747
Klaus Eder & Sarah M. Grundmann
ABSTRACT
The function of vitamin D in calcium homeostasis in dairy cows, such as in other vertebrates, is known for many years. In recent years, new and interesting, non-classical functions of vitamin D have been elucidated, including effects on the immune system. The major aim of this review is to provide an overview of effects of vitamin D or its metabolites on the immune system in dairy cows. The first part of the review provides an overview of vitamin D metabolism, with particular reference to the role of various proteins (25- and 1-hydroxylases, vitamin D binding protein, vitamin D receptor) in vitamin D signalling. The second part deals with the role of the concentration of 25-hydroxyvitamin D [25(OH)D] in plasma as an indicator of the vitamin D status in dairy cows, and its dependence on sunlight exposure and dietary vitamin D supplementation. In this part also the “free hormone hypothesis" is discussed, indicating that the concentration of free 25(OH)D might be a more valid indicator of the vitamin D status than the concentration of total 25(OH)D. The third part deals with classical and the non-classical functions of vitamin D. Among the non- classical functions which are based on an autocrine vitamin D signalling, particular reference is given to the effects of vitamin D and vitamin D metabolites on the immune system in bovine immune cells and in dairy cows. Recent findings provide some indication that vitamin D or its metabolite 25(OH)D could enhance the immune function in dairy cows and be useful for the prevention and therapy of mastitis. However, the number of studies reported so far in this respect is very limited. Thus, much more research is required to yield clear concepts for an optimised usage of vitamin D to improve the immune system and prevent infectious diseases in dairy cows.
📄 Download the PDF from VitaminDWiki
Introduction
The essential role of vitamin D in the homoeostasis of calcium and phosphorus in the body of animals has been well established for many years. For at least 25 years, it is known that vitamin D besides its classical functions exerts other functions which are not related to the calcium and phosphorus homoeostasis (Sassi et al. 2018). Among them, one very prominent function is the involvement of vitamin D in the immune system (Peelen et al. 2011; Ismailova and White 2021). In dairy cows, the immune system is suppressed during the peripartum phase which favours the occurrence of infectious diseases such as mastitis (Sordillo 2016). Several studies, mainly performed with human and murine immune cells, have shown that vitamin D stimulates the capacity of the innate immune system in the protection against pathogens (Martens et al. 2020; Ismailova and White 2021). These studies indicate that vitamin D could also exert beneficial effects in the prevention against infectious diseases in dairy cattle. For about 10 years, the role of vitamin D in the immune system of cattle is investigated, with a focus of vitamin D or its metabolites on the prevention or therapy of mastitis. The primary aim of this review is to provide an overview of potential effects of vitamin D or its metabolites on the immune system in dairy cows. The first section provides an insight into the complex metabolism of vitamin D, including absorption from the diet, activation by hydroxylation steps, and exertion of its function by binding to the vitamin D receptor ( VDR ). In the second section, the influence of vitamin D supply and exposure to sunlight on plasma concentration of 25- hydroxyvitamin D [25(OH)D], regarded as an indicator of vitamin D status, in dairy cows is presented. In this section, also the relevance of free 25(OH)D (which is not bound to proteins) in vitamin D signalling is discussed. In the third section, representing the main part, a brief introduction into the classical and non-classical functions of vitamin D is given, followed by a description of the actual knowledge of the effects of vitamin D on the innate and the adaptive immune system, which is mainly based on studies with human and murine immune cells. The final section deals with the studies performed with cattle or bovine cells to investigate the effects of vitamin D or vitamin D metabolites on the immune system and the pathogenesis of mastitis in cows.
Metabolism of vitamin D
Vitamin D exists in two different forms, vitamin D3 (cholecalciferol) which is of animal origin and vitamin D2 (ergocalciferol) which is formed in fungi and some plants (Japelt and Jakobsen 2013; Makris et al. 2020). Vitamins D2 and D3 share essentially the same metabolism, with the exception that they have different affinities for some proteins involved in vitamin D metabolism such as the vitamin D binding protein ( DBP ) and 25- hydroxylases, as discussed subsequently.
Vitamin D3 can be synthesised in the skin of most mammals (but not in dogs and cats) by a photochemical transformation of its precursor 7-dehydrocholesterol (7-DHC) (Zafalon et al. 2020). Vitamin D3 which is synthesised in the skin is released into the blood and bound to DBP which represents the main transport form of vitamin D in the blood (Duchow et al. 2019). Vitamin D as a part of the diet is taken up by active transport in the small intestine, with various proteins involved in lipid and cholesterol absorption [Scavenger receptor class B member 1, cluster of differentiation (CD) 36, Niemann-Pick C1 like intracellular cholesterol transporter 1] as transporters (Reboul et al. 2011). According to studies in mice, the rate of absorption might be similar for vitamins D2 and D3 (Baur et al. 2020). Interestingly, recent studies have shown that vitamin D is not only taken up into the intestinal epithelial cell but it is also excreted from the cell into the lumen. The efflux of vitamin D from the cell into the lumen is catalysed by ATP binding cassette subfamily B member 1 (ABCB1), a member of the ABC transporter family. It has been shown that the efflux of vitamin D from the cell into the lumen is critical for plasma concentrations of vitamin D3 and 25(OH)D, a vitamin D metabolite regarded as an indicator of the vitamin D status. ABCBl-knockout-mice showed plasma concentrations which were clearly in excess of those observed in their wild-type counterparts. Moreover, a single nucleotide polymorphism (SNP) in the ABCB1 gene observed in young men was associated with a reduction of 25(OH)D plasma concentration by 36% (Margier et al. 2019). In cattle, a potential role of the ABCB1 gene on the absorption of vitamin D has not been considered. However, it would be interesting to investigate the potential presence of SNPs in the ABCB1 gene of cattle which could be associated with changes in the efflux of vitamin D from intestinal cells.
In cattle, vitamin D originating from the diet will be exposed to the microbial community in the rumen before entering the site of absorption. Studies in the 70-ties and 80-ties of the past century suggested that a large part of vitamin D is decomposed in the rumen to at least four different biologically inactive compounds (Sommerfeldt et al. 1979, 1983). In vitro studies using rumen fluid revealed that around 80% of vitamin D is decomposed by microorganisms during a 24-h incubation period (Sommerfeldt et al. 1979; Horst and Reinhardt 1983). It has been suggested that the degradation of vitamin D in the rumen has the role to protect ruminants against toxic concentrations of vitamin D (Littledike and Horst 1982). Opposite to these previous findings, Hym0ller and Jensen (2010a) did not observe a loss of vitamin D in the rumen of cows equipped with a rumen fistula. Thus, the fate of vitamin D in the rumen is unclear. However, if a substantial loss of vitamin D in the rumen by microbial decomposition occurs, it is expected that the concentration of 25(OH)D in plasma resulting from a given vitamin D intake would be lower in ruminants than in monogastric species. In dairy cows, dietary intake of 20,000 IU/d, equivalent to about 30 IU/kg body weight (BW), resulted in average 25(OH)D plasma concentrations of 42 ng/ml (Nelson et al. 2016). In humans, an equivalent daily supply with vitamin D (2000 IU/d, representing about 30 IU/kg BW ), resulted in a very similar 25(OH)D plasma concentration of 41 ng/ml (Ekwaru et al. 2014). Cows of 12 practical herds with an intake between 30,000 and 50,000 IU/d (38,000 IU on average, equivalent to around 58 IU/kg BW) had an average 25(OH)D plasma concentration of 70 ng/ml, whereas humans with an equivalent supply (4000 IU/d, 61 IU/kg BW) had 25 (OH)D plasma concentrations of 50 ng/ml (Ekwaru et al. 2014). Although the metabolism of vitamin D between different species cannot be directly compared (i.e. rate of absorption, activity of 25-hydroxylases in the liver), the fact that cows responded similar or even better in the formation of 25(OH)D to a given amount of dietary vitamin D than humans, argues against a substantial decomposition of vitamin D by ruminal microorganisms in vivo.
Vitamin D, which is absorbed in the small intestine, is incorporated into chylomicrons in the epithelial cell. Chylomicrons are secreted from the epithelial cell into the lymph and enter the venous blood. A part of the vitamin D is taken up from chylomicrons into the liver while another part diffuses from chylomicrons to DBP (Dueland et al. 1983, 1985). Vitamin D is not biological active until it is hydroxylated two times at carbon-25 and carbon-1. The first hydroxylation step occurs in the liver and is catalysed by a group of 25-hydroxylases (CYP2R1, CYP27A1, CYP3A4, CYP2J2), with the microsomal P450 fraction localised CYP2R1 as the most important one (Jones et al. 2014a). CYP2R1 converts both, vitamins D3 and D2 into their 25-hydroxy derivatives; CYP27A1 prefers vitamin D3 as a substrate, while CYP3A4 and CYP2J2 prefer vitamin D2 as a substrate for 25-hydroxylation (Aiba et al. 2006; Baur et al. 2020). The efficiency of 25-hydroxylation could be different for vitamins D2 and D3. In humans, supplementation of vitamin D3 yielded higher concentrations of total 25-hydroxylated derivatives than supplementation of the identical amount vitamin D2 (Heaney et al. 2011; Lehmann et al. 2013). One reason for this might be that supplementation of vitamin D2 leads to a substantial decrease of the concentration of 25(OH)D3 (Lehmann et al. 2013). Moreover, it has been suggested that a lower concentration of total 25(OH)D in plasma after supplementation of vitamin D2 in comparison to vitamin D3 could be due to the shorter half-life of 25(OH)D2 in contrast to 25(OH)D3 (Heaney et al. 2011). Opposite to humans, mice supplemented with vitamin D2 showed a higher plasma concentration of total 25(OH)D than mice supplemented with vitamin D3 (Baur et al. 2020). It has been suggested that this observation in mice could be due to differences of either formation or degradation of the two different forms of 25(OH)D compared to humans (Baur et al. 2020). 25-hydroxylases are not regulated by the concentration of the substrate. However, it has been shown that the activity of CYP2R1 in mice is reduced in ageing and by obesity as well as by feeding a high fat diet (Roizen et al. 2018a, 2019). In humans, the formation of 25(OH)D from vitamin D is also reduced by obesity (Ekwaru et al. 2014; Carlsson et al. 2017). Moreover, in rat and mouse models, fasting caused a strong reduction of the expression of CYP2R1 in the liver (Aatsinki et al. 2019). In young goats, feeding a low protein diet increased the activity of CYP2R1 and plasma concentrations of 25(OH)D (Wilkens et al. 2019). In dairy cows, a SNP in the CYP2R1 gene, localised on chromosome 15, was associated with an increased incidence of mastitis (Naderi et al. 2018). This finding suggests that formation of 25(OH)D might be relevant with respect to development of mastitis. In cattle, there are no studies dealing with the effects of age or nutrition on the activity of CYP2R1 so far, but it would be interesting to investigate a possible effect of the body condition or the negative energy balance during early lactation on the formation of 25(OH)D. A small part of 25(OH)D formed in the liver (about 5%) is excreted via the bile while the large part is released into the circulation and bound to DBP . The complex of DBP and 25(OH) D is filtered by the glomerulus and reabsorbed by proximal tubule cells. The reabsorption takes place by binding of the DBP -25(OH)D complex to the megalin/cubulin receptor at the surface of tubule cells and subsequent endocytosis of the complex. Within the tubule cell, 25(OH)D is released from the complex by lysosomal degradation and translocated to mitochondria (Nykjaer et al. 2001; Saito et al. 2007). Within the mitochondrion, 25(OH) D is hydroxylated to 1,25-dihydroxyvitamin D [1,25(OH)2D] which represents the biologically active metabolite of vitamin D responsible for most, if not all, of the biologic actions of vitamin D (Makris et al. 2020). This hydroxylation step is catalysed by CYP27B1, an enzyme of the CYP450 family. 1,25(OH)2D (also called “calcitriol”) formed by the action of CYP27B1 is released into the blood, and bound, like other forms of vitamin D, to DBP . However, the affinity of human DBP for binding of 1,25(OH)2D is 10 to 100-fold lower than the affinity for binding of 25(OH)D (Verboven et al. 2002). The hydroxylation of 25(OH)D by CYP27B1 in the kidney is tightly controlled. Hydroxylation is mainly controlled by parathyroid hormone (PTH), fibroblast growth factor (FGF) 23 and 1,25(OH)2D. PTH, whose plasma concentration is increased under conditions of a low plasma calcium concentration stimulates the hydroxylation by an activation of CYP27B1. In contrast, FGF23 whose concentration is increased by high plasma concentrations of phosphate, inhibits hydroxylation. 1,25(OH)2D lowers hydroxylation by a negative feedback inhibition of CYP27B1 (Meyer and Pike 2020). Recent studies suggest that CYP27B1 is also inhibited by leptin, probably by an increase of the concentration of FGF23 (Reid et al. 2018). Both, 25(OH)D and 1,25(OH)2D can be metabolised by further enzymatic reactions. Among the catabolic enzymes, the mitochondrial 24-hydroxylase (CYP24A1) represents the most important one which catalyses multiple hydroxylation reactions at carbons 24 and 23 of the side chain of both 25(OH)D and 1,25(OH)2D. Hydroxylation of 1,25(OH)2D at carbons 23 and 24 initiates pathways to produce the biologically inactive calcitroic acid or 1,25(OH)2D-26,23-lactone as end products which are excreted via bile or urine (Makris et al. 2020). The major function of CYP24A1 therefore is to protect the body against toxic concentrations of 1,25(OH)2 D. CYP24A1 which has been identified in many tissues expressing the VDR is reciprocally regulated to CYP27B1 (Meyer and Pike 2020). CYP24A1 is a direct target gene of VDR , and therefore it is highly inducible by 1,25(OH)2D. The activity of CYP24A1 is moreover increased by FGF23, while it is lowered by PTH and hypocalcaemia (Jones et al., 2014a). The reciprocal regulation of CYP27B1 and CYP24A1 helps the body to regulate the concentration of 1,25(OH)2D at a physiological level (Meyer and Pike 2020). Recently, CYP3A4 has been identified as another microsomal enzyme which is able to inactivate 1,25(OH)2D by converting it into 1,23,25(OH)3D. The role of this enzyme in the inactivation of 1,25(OH)2D has been proven by the finding that a mutation of the gene, associated with an increased enzymatic activity, led even to the occurrence of vitamin D-dependent rickets in two patients in their childhood (Roizen et al. 2018b). An overview of the metabolism of vitamin D, including the hydroxylation steps, is shown in Figure 1.
Recent studies have shown that skeletal muscle might play an important role in the maintenance of the vitamin D status (Abboud et al. 2013, 2017). Skeletal muscle cells are able to take up DBP from blood via the megalin-cubulin receptor and incorporate it into the cell cytoplasm. DBP present in skeletal muscle cells is able to bind 25(OH)D which diffuses from blood plasma into cells. When DBP in the muscle undergoes proteolytic breakdown, 25(OH)D is released and diffuses into the blood where it is available for binding to DBP (Rybchyn et al. 2020). Although the concentration of 25(OH)D in skeletal muscle is low (1-3 |ug/kg in sheep and cattle, Liu et al. 2013), the whole amount of 25(OH)D in muscle might be comparable to that in the circulation due to the great amount of muscle mass. It has been suggested that the uptake of 25(OH)D into the muscle and its release into the blood accounts for the very long half-life of this metabolite in the circulation (Rybchyn et al. 2020). Studies in myotubes have shown that the uptake and release of 25(OH)D is regulated by PTH (Abboud et al. 2017). In free living animals, a release of 25(OH)D from muscle into the blood might be important for maintaining a sufficient level of 25(OH)D in the blood during the winter season when vitamin D synthesis in the skin is lacking.
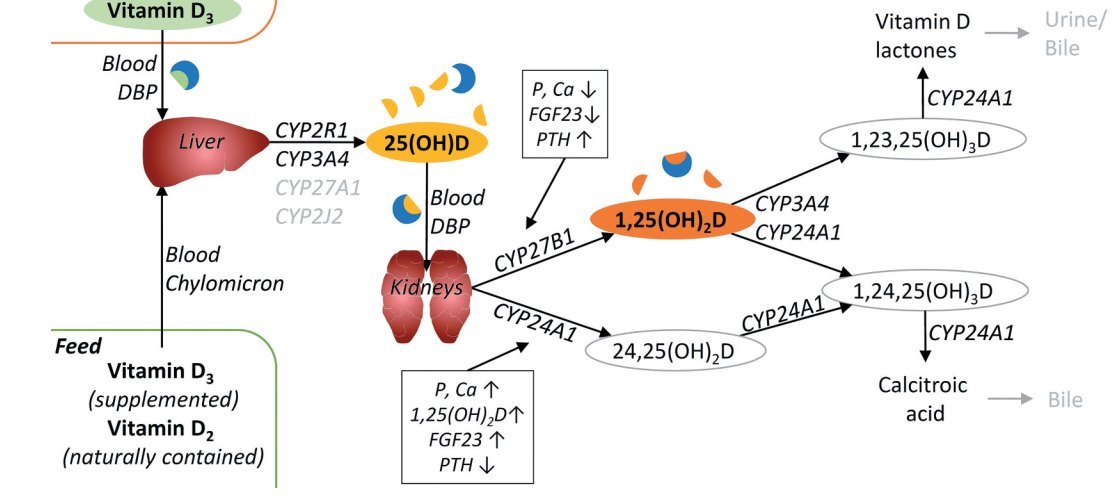
Figure 1. Metabolism of vitamin D.
Vitamin D synthetised in the skin or deriving from the feed is absorbed in the small intestine and delivered to the liver where it is hydroxylated to 25-hydroxyvitamin D [25(01-1)D] by various 25-hydroxylases (CYP2R1, CYP27A1, CYP3A4, CYP2J2). 25(OH)D is transported to the kidney, where it is hydroxylated by CYP27B1 to 1,25-dihydroxyvitamin D [1,25(OH)2D], the biological active metabolite of vitamin D. Both, 25(OH)D and 1,25(OH)2D are also hydroxylated, mainly by CYP24A1, to non-bioactive metabolites which are excreted via urine or bile. The activities of CYP27B1 and CYP24A1 are regulated by various factors [concentrations of phosphorus (P), calcium (Ca), fibroblast growth factor 23 (FGF23), parathyroid hormone (PTH), 1,25(OH)2D] in order to maintain the concentration of 1,25(OH)2Data physiological level. In the blood, the vitamin D metabolites are primarily bound to vitamin D binding protein ( DBP ), but a small percentage is also available in free form. Cells expressing the megalin/cubulin receptor are able to take up DBP -bound vitamin D metabolites [25(OH)D, 1,25(OH)2D] by endocytosis. In cells not expressing the megalin/cubulin receptor vitamin D metabolites in free (unbound) form are taken up by diffusion. Cells expressing CYP27B1 are able to form 1,25(OH)2 D from 25(OH)D. Within the cell 1,25(OH)2D binds to the vitamin D receptor ( VDR ), which dimerises with retinoid X receptor (RXR) and other cofactors. The VDR -RXR complex binds to vitamin D response element ( VDR E) of the DNA and mediates the effects of vitamin D by regulating the transcription of target genes. For more detail and references, see text.
The biological effects of 1,25(OH)2D are mediated by the vitamin D receptor ( VDR ). VDR , which is a member of the nuclear receptor superfamily of ligand- activated transcription factors, is widely expressed in tissues of the body. Binding of 1,25(OH)2D to VDR causes an activation of the receptor which in turn forms a heterodimeric complex with the retinoid X receptor (RXR). The VDR -RXR complex binds to the promotor regions of target genes, known as vitamin D response elements, and in interaction with nuclear coactivators and transcription factors induces the transcription, and/or suppression of genes. Gene expression studies in humans have shown that 1,25(OH)2D directly or indirectly regulates more than 2000 genes involved in many pathways including pathways which are not involved in calcium homoeostasis (Ramagopalan et al. 2010). One of the most prominent among these pathways is the involvement of vitamin D signalling in the regulation of both the innate and the adaptive immune system (Koivisto et al. 2020; Ismailova and White 2021). Besides 1,25(OH)2D which represents the physiological ligand of VDR , 25(OH)D can bind to VDR with a much lower affinity (Marcinowska- Suchowierska et al. 2018). While the concentration of 1,25(OH)2D in plasma is tightly controlled, formation of 25(OH)D is uncontrolled meaning that dietary supplementation of excess doses of vitamin D results in very high plasma concentrations of 25(OH)D. Plasma concentrations of 25(OH)D which are in strong excess of physiological concentrations can lead to an overstimulation of VDR , which as a result of the induction of a too strong vitamin D response, in turn causes symptoms of hypervitaminosis D which are induced by the development of hyper- calcaemia and calcinosis. In cattle, the main symptoms of calcinosis are emaciation, extended lying, locomotion disorders, raised pulse and respiratory rate, impaired fertility as well as decreased vitality, altogether resulting in high economic losses (Hodnik et al. 2020). In dairy cows, hypervitaminosis D can also be induced by ingestion of calcinogenic plants, such as Solanum malacoxylon, Cestrum diurnum, Trisetum flavescens and Nierembergia veitchii (Wasserman 1975; Mello 2003). These plants contain 1,25(OH)2D3 glycosides. In the rumen, 1,25(OH)2D is released from these glycosides by microbial cleavage of the sugar residue. A part of the released 1,25(OH)2D is absorbed, enters the blood stream and overcomes the physiological regulation of the concentration of 1,25(OH)2D. Unphysiological high concentrations of 1,25(OH)2D in plasma then lead to hypercalcaemia and calcinosis (Mello 2003).
Besides its effect on the transcription of genes, VDR can also exert non-genomic responses as a receptor located in sphingolipid and cholesterol-rich membranes (Huhtakangas et al. 2004). Binding of 1,25(OH)2D to this receptor leads to the activation of various signalling pathways, including guanylate cyclase, phosphatidyli- nositol-3-kinase (PIK3), cyclic adenosine monophosphate (cAMP), mitogen-activated protein kinase (MAPK), protein kinase (PK) A, and phospholipase C (PLC) pathways, the opening of voltage-dependent calcium and chloride channels and mobilisation of calcium from the sarcoplasmic reticulum (Zafalon et al. 2020). Non-genomic responses of 1,25(OH)2D mediated by membrane localised VDR occur rapidly over minutes or hours and include rapid intestinal absorption of calcium, secretion of insulin by pancreatic cells or migration of endothelial cells (Saponaro et al. 2020). In humans, various SNP of the VDR were identified which were associated with an increased risk for various diseases including various types of cancer (Yu et al. 2018; Gnagnarella et al. 2021), coronary artery disease (Tabaei et al. 2021), osteoarthritis (Li et al. 2021), osteoporosis (Wu et al. 2016) and others. Based on these findings, potential effects of polymorphisms in the VDR of cattle on vitamin D signalling would be of interest. Deiner et al. (2012) aimed to investigate the relationship between sequence alterations in the VDR with the incidence of periparturient hypocalcaemia in 26 dairy cows. The authors detected eight sequence alterations, with four of them located on exons thereby potentially causing changes of the encoded amino acid of the VDR protein. There were no associations between sequence alterations and the incidence of periparturient hypocalcaemia, which however could be due to the low number of animals considered.
Recent studies suggested that, besides VDR , other membrane-associated vitamin D-binding proteins are involved in the non-genomic rapid response to 1,25(OH)2 D (Zmijewski and Carlberg 2020). One of these candidates is PDIA3 (protein disulphide isomerase family A member 3), also known as 1,25(OH)2D-MARRS (membrane- associated rapid response to steroid), which has been shown to be involved in calcium uptake in intestinal cells (Nemere et al. 2012). A role of PDIA3 in calcium homoeostasis has been supported by the finding that PDIA3 knockout in mice was associated with abnormalities in skeletal tissue (Wang et al. 2010). In cattle, the role of PDIA3 has not yet been investigated. However, it would be of interest to investigate the relevance of PDIA3 on calcium homoeostasis in dairy cows, particularly during the periparturient phase, when cows are at the risk of hypocalcaemia.
Assessment of the vitamin D status of dairy cows
25(OH)D as an indicator of the vitamin D status
Although 1,25(OH)2D is the bioactive metabolite of vitamin D, its concentration is not an appropriate indicator of the vitamin D status as its concentration in plasma is tightly regulated. Moreover, 1,25(OH)2D has a short half-life of about 15 h (Chun et al. 2019). In contrast, the formation of 25(OH)D is not regulated, and its plasma concentration is strongly responding to the vitamin D intake. Moreover, the half-life of 25(OH)D, being in the range of 2-3 weeks, is much greater than that of 1,25(OH)2D. As an additional aspect, blood levels of 25(OH)D are much higher and easier to measure compared to 1,25(OH)2D. Therefore, plasma concentration of 25(OH)D is regarded as an appropriate indicator of vitamin D supply (Chun et al. 2019). 25(OH)D present in plasma is almost completely bound to proteins, whereas only a very small part exists in free form. In cattle, plasma concentrations of 25(OH)D below 5 ng/ml are regarded as an absolute deficiency of vitamin D leading to specific vitamin D deficiency symptoms such as rickets or osteomalacia (Horst et al. 1994). It has been suggested that plasma concentrations of 25(OH)D greater than 20 ng/ml in cattle are required for the maintenance of a physiologic homoeostasis of calcium and phosphorus (NRC 2001). Plasma 25(OH)D concentrations required for the enhancement of the immune system in dairy cows have not yet been investigated. Studies in humans indicate that plasma 25(OH)D concentrations >30 ng/ml are needed to support optimal immune function. Nelson et al. (2012) suggested that, given the similarities between humans and cattle regarding vitamin D physiology, a similar concentration of 25(OH)D might be required in cattle to provide an optimum function of the immune system. However, recent findings suggest that much higher, supraphysiological concentrations of 25(OH)D in plasma could be required to enhance the immune function in dairy cows (see chapter 5.2). Prolonged plasma concentrations of 25(OH)D greater than 200 ng/ml in cows are regarded as critical with respect to the development of hypervitaminosis D (Littledike and Horst 1982).
Vitamin D3 can be synthesised in the skin from 7-DHC as a precursor by exposure to sunlight at a wavelength between 290 and 315 nm. 7-DHC, which occurs in deep layers of the epidermis as a component of the membranes’ phospholipid bilayer, represents an intermediary product in the cholesterol synthesis pathway (Norman 2012). Therefore, vitamin D3 synthesis in the skin not only depends on the light intensity and wavelength but also on the concentration of 7-DHC in the skin which is converted by 7-DHC reductase into cholesterol. An increased activity of this enzyme leads to a reduction of the concentration of 7-DHC, and in turn to a reduced rate of vitamin D3 synthesis (Prabhu et al. 2017). The relationship between the activity of 7-DHC reductase, the concentration of 7-DHC in the skin and vitamin D3 synthesis could be also relevant in cows, but has not been investigated so far. In cattle, vitamin D3 synthesis takes place in all areas of the skin and is not exclusively associated with skin areas where hair coverage is scant or lacking such as in udder or muzzle (Hym0ller and Jensen 2010b). Vitamin D3 synthesis in the skin depends on light intensity and the duration of exposition. With increasing distance from the equator, the proportion of UV-B in the daylight spectrum and the period with a sufficient UV-B proportion for vitamin D3 synthesis decrease. Without dietary vitamin D supply and lack of exposition to UV-B light, plasma concentrations of 25(OH)D in plasma are strongly decreasing. In the study of Hym0ller et al. (2017), plasma concentrations of 25(OH)D after a 6-month period without vitamin D-supplementation and daylight exposition fell to values below 5 ng/ml, representing a deficiency of vitamin D. Exposition of the cows to UV-light from a UV lamp for 2 h per day over a period of 28 d without additional dietary vitamin D supplementation led to an increase of plasma 25(OH)D concentration from <5 to about 35 ng/ml (Hym0ller et al. 2017). This study indicated that vitamin D3-deprived cattle are able to increase their plasma 25(OH)D concentrations by a maximum of 1 ng/ml per day by UV-light exposure. In the study of Hym0ller and Jensen (2012), a daily pasture time of 5 h at a latitude degree of 56°N (Denmark) raised 25(OH)D plasma concentration from 22 ng/ ml to 37 ng/ml during a period of 28 d without an additional supply of vitamin D. At a daily pasture time of 2.5 h, plasma 25(OH)D concentration rose only slightly (from 20 to 23 ng/ml), at a daily pasture time of 1.25 h, plasma 25(OH)D concentration remained at a constant level, and at a daily pasture time of only 0.5 h, plasma 25(OH)D concentration fall to a value of about 10 ng/ml. During the summer months, if there is sufficient UV-B exposure, an additional oral supply of vitamin D causes no or at least a moderate increase of plasma 25(OH)D concentrations (Hidiroglou et al. 1979; Hym0ller et al. 2009). In the study of Hym0ller et al. (2009), cows kept on pasture from May to October had plasma 25(OH)D concentrations of about 40 ng/ml during July and August. Additional daily supplementation of 20,000 IU vitamin D3 increased plasma 25(OH)D concentration by 5 ng/ml to about 45 ng/ml during these months.
If exposition to sunlight is lacking, plasma concentration of 25(OH)D is mainly depending on the dietary vitamin D supply. NRC (2001) recommends a dietary vitamin D intake of 30 IU/kg BW, according to a total vitamin D supply of 20,400 IU for a cow with a BW of 680 kg. In a practical cow herd supplemented with 20,000 IU per cow and day, average plasma concentrations of 25(OH)D were about 42 ng/ml. However, 22% of the cows of this herd had plasma 25(OH)D concentrations below 30 ng/ml. In the same study, cows from 11 other herds from various federal states of the USA were considered which received vitamin D in daily amounts between 30,000 und 50,000 IU/d, representing the common practical vitamin D supply of dairy cows in the USA. These cows showed an average 25 (OH)D plasma concentration of 70 ng/ml (Nelson et al. 2016). In some studies, the effect of oral supplementation of 25-hydroxyvitamin D3 [25(OH)D3] on vitamin D status was investigated. These studies show that supplementation of 25(OH)D3 causes a much stronger increase of 25(OH)D plasma concentration in cows than supplementation of the equivalent amount of vitamin D3. In the study of Wilkens et al. (2013) daily supplementation of 3 mg 25(OH)D3 caused an increase of the plasma concentration of 25(OH)D from about 30 ng/ml to about 120 ng/ml in dairy cows within a short period of 6 d. Interestingly, the incline of the 25(OH)D concentration in this study was greater in cows of the second parity than in cows of the third parity, and the half-life after discontinuation of the supply with 25(OH)D was greater in second parity cows than in third parity cows. This study indicates that the age influences degradation of 25(OH)D in cows. In the study of Poindexter et al. (2020), oral supplementation of 1 mg or 3 mg vitamin D3 (equivalent to 40,000 or 120,000 IU, respectively) per day resulted in 25(OH)D plasma concentrations of 57 or 77 ng/ml, resp., after an 8-week period. Oral supplementation of the same amounts (1 or 3 mg) of 25(OH)D3 resulted in plasma concentrations of 180 or 278 ng/ml, resp., after a 8-week period. In contrast, the resulting plasma concentration of 1,25(OH)2D was similar within the groups receiving either vitamin D3 or 25(OH)D3, which confirms the observation that the synthesis of 1,25(OH)2D is tightly controlled. Concentrations of calcium and phosphorus remained unchanged by supplementation of 25(OH)D3, indicating that even the high 25(OH)D concentrations observed in the group supplemented with 3 mg 25(OH)D3, which were clear in excess of 200 ng/ml , did not cause a hypervitaminosis, at least during the 8-week period considered. Similar results were obtained in a study in which cows received a daily oral dose of 2 mg 25(OH)D3 from day 21 prepartum until the day of partum, and/or a daily dose of 1 mg 25(OH)D3 from the day of partum until day 21 in lactation (Golder et al. 2021). In this study, plasma 25(OH)D concentrations in the group supplemented during the prepartum and the postpartum period rose to a value of 239 ng/ ml, without significantly affecting plasma calcium concentration. In the study of Rodney et al. (2018), cows received an oral dose of 3 mg 25(OH)D3 from day 255 of gestation until the day of partum as a part of a diet with either a positive or a negative dietary cation anion difference (DCAD). This study showed that DCAD can influence vitamin D metabolism. While the cows in the group fed the diet with a positive DCAD showed a 25(OH)D concentration of about 234 ng/ml, the group fed the diet with a negative DCAD showed a significantly lower 25(OH)D plasma concentration of about 203 ng/ml. In this study, concentrations of free and total calcium were slightly increased by 25(OH)D3 supplementation but remained within the physiological range. In this study, daily supplementation of 3 mg (120,000 IU) vitamin D3 resulted in plasma 25(OH)D concentrations of 60 ng/ml. This finding confirms that plasma 25(OH)D concentrations reached by vitamin D3 supplementation are much lower than those reached by supplementation of 25(OH)D3. The reason for the effect of DCAB on 25(OH)D concentration has not been elucidated so far.
Plasma 25(OH)D concentrations are also influenced by the stage of lactation. It has been shown that plasma 25(OH)D concentrations in cow herds are lower at early lactation than at later lactation or during the dry period, even if the daily vitamin D3 intake is similar during the different periods (Nelson et al. 2016; Holcombe et al. 2018). It has been well established that cows are undergoing a systemic inflammation during early lactation (Gessner et al. 2013; Bradford et al. 2015). Although there is no direct evidence in dairy cows, it is well known that inflammatory challenges (such as resulting from surgeries) or chronic diseases associated with the occurrence of inflammation in humans are associated with a decrease of plasma 25(OH)D concentration (Louw et al. 1992; Guillot et al. 2010; Reid et al. 2011; Garbossa and Folli 2017), and plasma 25(OH)D concentration has even been regarded as a negative acute-phase reactant (Waldron et al. 2013). It has been hypothesised that inflammation enhances the formation of 1,25(OH)2 D in extrarenal tissues by an activation of CYP27B1 which in turn inhibits the formation of 25(OH)D from vitamin D in the liver (Mangin et al. 2014). Therefore, it is likely that reduced plasma concentrations of 25(OH)D in cows during early lactation are due to a reduced formation of 25(OH)D in the liver as a consequence of a dysregulation of the vitamin D status due to various causes of inflammation such as metabolic stress or infectious diseases.
Role of free 25(OH)D in vitamin D signalling
As discussed in the previous chapter, plasma concentration of 25(OH)D is regarded as an appropriate indicator of vitamin D status. In plasma, 25(OH)D formed in the liver by hydroxylation of vitamin D is bound with high affinity to DBP and with low affinity to albumin. In humans, 85-88% of 25(OH)D is bound to DBP and 12-15% is bound to albumin (Safadi et al. 1999). The concentration of DBP is in large excess in relation to the concentration of vitamin D metabolites. Therefore, only a small part of DBP (<2%) exists as holoprotein ( DBP plus vitamin D-metabolite) (Bouillon et al. 2020). 25(OH)D bound to DBP can be taken up into the tubule cell in the kidney via the megalin/cubulin receptor. In the kidney, a small part of 25(OH)D is then converted into 1,25(OH)2D and released into the blood. Plasma concentrations of 25(OH)D and 1,25(OH)2D in cattle are similar to those in humans. In both species, the concentration of 25(OH)D in plasma is about 1000-fold higher than that of 1,25(OH)2D (Rodney et al. 2018; Chun et al. 2019; Bouillon et al. 2020; Poindexter et al. 2020). 1,25(OH)2D present in the circulation originates exclusively from the formation in the kidney. Besides the kidney, many other cells in the body express both, CYP27B1 and VDR (see chapter 4.2). These cells are able to form 1,25(OH)2D from 25(OH)D, which enables them to an autocrine/intracrine vitamin D-signalling. However, as most of these cells with the exception of cells from parathyroid gland, placenta and skeletal muscle do not express the megalin/ cubulin receptor at their surface, these cells are not able to internalise the DBP -25(OH)D complex by endocytosis (Bikle 2020). Therefore, 25(OH)D required for the production of 1,25(OH)2D is taken up into these cells in its free form by diffusion (Chun et al. 2019). Due to its high binding affinity to DBP , the concentration of free 25(OH)D is very low (Bouillon et al. 2020). In humans, the concentration of free 25(OH)D lies in the range of 4-8 pg/ml, representing less than 0.05% of the total 25(OH)D (Bikle et al. 1986; Chun et al. 2019). In cattle, the concentration of free 25(OH)D has not been reported so far, probably as the analysis of free 25(OH)D in plasma, due to its low concentrations, is challenging. However, binding kinetic studies have shown that DBP of cattle has even a higher binding affinity to 25(OH)D than human DBP (Kd, 1.96 vs. 7.54 nM), and the ratio between free and total 25(OH)D is about 3-fold lower in cattle than in humans (0.55 vs. 1.50 pmol free 25(OH)D/nmol total 25 (OH)D) (Vieth et al. 1990). Although the concentration of free 25(OH)D in plasma is very low, this fraction might be important as a substrate for the production of 1,25(OH)2D in cells which are not able to internalise 25(OH)D bound to DBP . Experiments in animal models support the hypothesis that vitamin D metabolites in free form play an important role in vitamin D signalling. Two studies with mice have shown that deletion of the DBP gene leads to strongly reduced plasma concentrations of total 25(OH)D and 1,25(OH)2D (Safadi et al. 1999; Zella et al. 2008). However, calcium homoeostasis was not affected in these mice in comparison to their wildtype counterparts. Moreover, in spite of the largely reduced concentrations of total 25(OH)D in plasma, concentrations of vitamin D metabolites in tissues were not lowered in mice with DBP deletion (Zella et al. 2008). Distribution, uptake, activation profile and biologic potency of 1,25(OH)2D after injection of 1,25(OH)2D were also not different between the DBP knockout genotype and the wildtype genotype (Zella et al. 2008). However, depletion of vitamin D led to a faster disturbance of calcium homoeostasis (hyperparathyroidism, bone mineralisation defects) in the DBP knockout genotype than in the wildtype genotype. On the other hand, DBP knockout mice were more protected against vitamin D toxicity by exposure to high doses of vitamin D, probably by a more rapid metabolism of vitamin D metabolites than mice of the wildtype genotype (Safadi et al. 1999). These studies indicate that the major role of DBP is to maintain a stable store of vitamin D metabolites and to regulate their free and total levels in plasma (Bikle and Schwartz 2019). Recently, the case of a 58-year old woman with a homozygous deletion of DBP has been reported. Although this woman had very low concentrations of 25(OH)D and 1,25(OH)2D in plasma, concentrations of calcium and PTH were in the physiologic range, and there was no history of rickets or signs of osteomalacia (Henderson et al. 2019). This case report supports the view that functions of vitamin D metabolites are independent of their binding to DBP .
There are also some studies in humans which indicate that concentrations of free 25(OH)D in plasma are more closely related to vitamin D associated parameters than total 25(OH)D. A study in healthy young adults found that concentrations of free and bioavailable (free + albumin-bound) 25(OH)D show a higher correlation with bone mineral density than total 25(OH)D (Powe et al. 2011). In several studies with human participants supplied with vitamin D, plasma concentration of free 25(OH)D was inversely correlated with PTH concentrations, while total 25(OH)D was not (Schwartz et al. 2016; Shieh et al. 2016, 2018; Wang et al. 2017). The relevance of free 25(OH)D for vitamin D signalling in cells expressing CYP27B1 is also supported by the fact that deficiency symptoms of vitamin D are occurring solely under the condition of a low plasma 25(OH)D concentration while plasma concentration of 1,25(OH)2D is typically normal or even increased under such conditions, as a result of an activation of CYP27B1 in the kidney (Chun et al. 2019).
The free hormone hypothesis postulates that only the non-bound (free) fraction of a hormone is able to enter the cell and exert its biological function. This hypothesis applies, for example, to thyroid hormones, sex steroids and cortisol (Bikle 2020). According to the findings mentioned above, there are several strong arguments which indicate that the free hormone hypothesis applies also to vitamin D metabolites, at least for the non-classical vitamin D functions. A more detailed discussion of the potential role of free 25(OH)D for vitamin D signalling in cells not expressing the megalin/cubulin transport system has been published recently in several reviews (Bikle et al. 2017a, 2017b; Chun et al. 2019; Bikle 2020). In cattle, the potential role of free 25(OH)D in vitamin D signalling has not yet been investigated. However, based on findings in humans and mice, it is highly probable that the concentration of free 25(OH)D in plasma might also be important in cattle for vitamin D signalling in cells which do not express the megalin/ cubuline transport system such as cells of the immune system. With respect to plasma concentration of free 25(OH)D, plasma concentration and protein structure of DBP might be also of relevance which as a binding protein influences the concentration of free 25(OH)D. In humans, DBP is genetically very polymorphic with three frequent alleles, and six major phenotypes (Bouillon et al. 2020). It has been shown that these phenotypes differ in their affinity for vitamin D metabolites (Arnaud and Constans 1993) and that the DBP haplotype has significant effects on total 25(OH)D, free 25(OH)D, and DBP concentrations (Schwartz et al. 2018). In dairy cows, DBP has been identified as strong candidate gene associated with incidence of clinical mastitis (Abdel-Shafy et al. 2018). These findings clearly imply that DBP is a key component in vitamin D signalling by influencing the concentrations of free vitamin D metabolites in plasma. Another point relevant for dairy cows is the observation that DBP is able to bind fatty acids, particularly unsaturated fatty acids (oleic acid, linoleic acid and arachidonic acid) (Williams et al. 1988). Mono- and polyunsaturated fatty acids impair the binding of 25(OH)D and 1,25(OH)2D to DBP . It has been shown that concentrations of 36 uM of linoleic acid or arachidonic acid decrease the affinity of DBP to vitamin D metabolites to 22 to 42% (Bouillon et al. 1992). In dairy cows, concentrations of non-esterified fatty acids (NEFA) in plasma are several-fold higher than 36 uM, in particular during the first weeks of lactation, at which NEFA typically reach levels of 0.5 to 1 mM (Schlegel et al. 2012; Gessner et al. 2015). The plasma NEFA fraction consists largely of saturated fatty acids (up to 90%), while the proportions of unsaturated fatty acid are relatively low (up to 10%, with oleic acid as the main component) (Contreras et al. 2010; Liu et al. 2020). In spite of the relatively low proportions of unsaturated fatty acids in the total plasma NEFA fraction, their absolute concentrations in plasma could be sufficient to suppress the binding of vitamin D metabolites to DBP , including 25(OH)D, and thus lead to increased concentrations of vitamin D metabolites in their free form. An increased concentration of free 25(OH)D then could stimulate vitamin D signalling in cells not expressing the megalin/cubulin transporter. Although this matter is speculative, it would be worth of being investigated in cattle.
Previous studies have shown that DBP has a lower binding affinity to 25(OH)D2 than to 25(OH)D3 (Jones et al. 1980; Hollis 1984). This finding not only explains the fact that 25(OH)D2 has a shorter half-life than 25(OH)D3 (Jones et al. 2014b) but also indicates that supplementation of vitamin D2 might yield a higher concentration of its free 25- hydroxy metabolite than supplementation of the same amount of vitamin D3. Indeed, concentrations of free 25(OH)D2 in plasma of mice supplemented with vitamin D2 were two- to three-fold higher than the concentrations of free 25(OH)D3 in mice receiving the identical amount of vitamin D3 (Chun et al. 2016; Baur et al. 2020). In contrast, plasma concentrations of 1,25(OH)2D2 in these studies were lower in the mice supplemented with vitamin D2 than that of 1,25(OH)2D3 in mice supplemented with vitamin D3. A potential explanation for this finding is that the lower binding affinity of DBP for 25(OH)D2 relative to 25(OH)D3 may impair the availability of the 25(OH)D2- DBP complex for megalin/cubulin-mediated glomerular reabsorption (Chun et al. 2016; Baur et al. 2020). The observation that supplementation of vitamin D2 yields a higher concentration of the free 25-hydroxy-vitamin D2 metabolite in comparison to supplementation of vitamin D3 suggests that supplementation of vitamin D2 could enhance the autocrine vitamin D signalling in cells which take up free 25(OH)D from plasma as a substrate for formation of the 1,25(OH)2D. Such an autocrine vitamin D signalling occurs predominately in cells which are involved in the non-classical, extraskeletal functions of vitamin D, including the immune system. On the other hand, reduction of the concentration of the 1,25-dihydroxy metabolite in plasma by supplementation of vitamin D2 in comparison to vitamin D3 suggests that the classical vitamin D functions which are based on 1,25(OH)2D released from the kidney are weakened by vitamin D2 in relation to vitamin D3. Thus, overall, it would be expected that vitamin D2 shifts vitamin D signalling from classical (i.e. skeletal) to non-classical (i.e. extraskeletal) functions of vitamin D in comparison to vitamin D3.
Functions of vitamin D
Classical functions of vitamin D
In all vertebrates, the classical function of vitamin D is to regulate calcium and phosphorus homoeostasis and bone metabolism. This function is well established for many years, and therefore will be only briefly described in this review.
The classical target tissues of vitamin D involved in the regulation of calcium and phosphorus homoeostasis are small intestine, kidneys, parathyroid gland and bone. In the small intestine, 1,25(OH)2D stimulates the transcellular and the passive paracellular calcium transport. The transcellular transport of calcium is stimulated by an increased expression of transient receptor potential vanilloid type 6 (TRPV6), which - located in the apical membrane - facilitates the entry of calcium in the cell, an upregulation of calbindin-D9k which facilitates the transfer across the cell, and an upregulation of Ca2+-ATPase which extrudes calcium through the basolateral membrane into the circulation (Christakos et al. 2011). 1,25(OH)2D moreover stimulates paracellular transport of calcium by increasing the permeability of tight junctions due to a suppression of the expression of various tight junction proteins (Kutuzova and Deluca 2004). 1,25(OH)2D also enhances the absorption of phosphate by an upregulation of the Na-Pi-transporter (Kiela and Ghishan 2016). In ruminants, a part of the calcium absorption takes places in the rumen. The pre-duodenal net absorption of calcium in cattle depends on the calcium intake. At dietary intakes of calcium below 120 g/d in cattle, the pre-duodenal calcium net absorption rate is very low. If this dietary calcium intake level is exceeded, the pre-duodenal calcium net absorption rate is strongly increasing. At the same time, an increase of the pre-duodenal calcium absorption is associated with a reduction of the intestinal calcium net absorption rate, meaning that the pre-duodenal absorption of calcium becomes more important for whole calcium absorption at high dietary calcium intake (Schröder and Breves 2006). However, the pre-duodenal absorption of calcium is probably not mediated by classical mechanisms as TRPV6 and calbindin-D9k are lacking in rumen epithelial cells, and thus is not driven by vitamin D (Wilkens et al. 2009; Wilkens and MuscherBanse 2020).
In the kidney, 1,25(OH)2D stimulates calcium reabsorption by enhancing the expression of TRPV5, calbindin-D28K protein and the basolateral calcium transporters Ca2+- ATPase (PMCA1b) and Na+, Ca2+ exchanger (NCX) at distal and connecting tubules (Wilkens and Muscher-Banse 2020). TRPV5 which is located at the apical membrane represents an epithelial calcium channel which enables the transport from primary urine into the tubular cell. Calbindin-D28K facilitates the transport of calcium from the apical to the basolateral side of the cell where calcium is released into the blood by the two calcium transporters, PMCAlb and NCX (Boros et al. 2009). Renal tubular reabsorption of phosphate is stimulated by 1,25(OH)2D by decreasing the PTH level (Jacquillet and Unwin 2019).
In the parathyroid gland, 1,25(OH)2D decreases gene transcription of PTH (Silver et al. 1985). The reduction of PTH synthesis is due to a suppression of the parathyroid gland cell growth caused by a decreased production of growth factors (transforming growth-factor-epidermal growth factor receptor growth loop), and by an increased production of inhibitors of cell growth (cyclin-dependent kinase inhibitors p21 and p27) (Dittmer and Thompson 2011). Moreover, 1,25(OH)2D induces an upregulation of the calcium-sensing receptor in the parathyroid gland, which makes the gland more sensitive to plasma ionised calcium, which in turn leads to a decreased PTH secretion (Zafalon et al. 2020).
The effect of 1,25(OH)2D on bone tissue depends on plasma calcium concentration. In a hypocalcemic condition, 1,25(OH)2D in conjunction with PTH stimulates mobilisation of calcium from bones following the aim to increase plasma concentration of ionised calcium (Baldock et al. 2006). This effect of 1,25(OH)2D is caused by a stimulation of the activator receptor for nuclear factor-kappa B (NF-kB) ligand (RANKL) which increases the formation and the activity of osteoclasts (Anderson and Atkins 2008). 1,25(OH)2 D moreover lowers the production of osteoprotegerin, a protein which inhibits osteo- clastogenesis, and in turn also stimulates bone resorption (Boyce and Xing 2008). Under conditions of a positive calcium balance, 1,25(OH)2D stimulates bone growth and mineralisation. This effect is caused indirectly, by increasing calcium and phosphate concentrations in plasma, and by direct effects exerted in osteoblasts (Zafalon et al. 2020). Osteoblasts express VDR , and 1,25(OH)2D directly affects their growth and differentiation by regulating the expression of differentiation and mineralisation-related genes such as alkaline phosphatase, osteocalcin and osteopontin (van de Peppel and van Leeuwen 2014).
Non-classical functions of vitamin D
For about 30 years, it is known that not only classical vitamin D target tissues, but also most other tissues express a functional VDR . Moreover, many tissues also express CYP27B1 meaning that they are able to produce 1,25(OH)2D (Bikle 2009; Bouillon and Bikle 2019). Cells expressing both, VDR and CYP27B1 are able to perform an autocrine/ intracrine vitamin D signalling. An overview of cells enabled to co-express a functional CYP27B1 and VDR , according to Chun et al. (2019), is shown in Table 1. 1,25(OH)2 D formed in these cells is not released into the circulation but is retained for local actions (Meyer and Pike 2020). In most of these cells CYP27B1 is regulated differently from CYP27B1 in the kidney. For instance, CYP27B1 in macrophages and monocytes is activated by immune activators while it is not feedback inhibited by 1,25(OH)2D. In keratinocytes, CYP27B1 expression is also stimulated by immune activators including tumour necrosis factor-a (TNFa) and interferon-y (IFN-y), while in bone cells it is!!!!!Table 1. Cell types
capable of autocrine vitamin D signalling by co-expressing CYP27B1 and VDR (Chun et al. 2019). stimulated by PTH and 25(OH)D (Bouillon and Bikle 2019). As a result of autocrine/ intracrine vitamin D signalling, vitamin D exerts various effects which are not related to calcium and phosphate homoeostasis. The actions of vitamin D include effects on the immune system, effects on heart and cardiovascular system, effects on adipose tissue and glucose/lipid metabolism, effects on muscle and effects related to the pathogenesis and progression of cancer (Saponaro et al. 2020). As dairy cows are very prone to various infection diseases such as mastitis during the periparturient period, the immune system is of extraordinary relevance during this phase. Therefore, in the following sections, this review will deal with the role of vitamin D in the immune system.
The role of vitamin D in the immune system
For at least two decades, it is known that vitamin D exerts important functions in both, the innate and the adaptive immune system (Bikle 2008; Peelen et al. 2011; Martens et al. 2020; Ismailova and White 2021). Most of the knowledge in this respect has been gained from in vitro studies with human or murine immune cells. In the present section, effects of vitamin D on the immune system observed in these studies are briefly described.
Effects of vitamin D in the innate and the adaptive immune system
Monocytes und macrophages which are key players in the innate immune system express CYP27B1 and thus are able to produce 1,25(OH)2D from 25(OH)D which enters the cell from the blood. As they express an active form of VDR , vitamin D signalling in an autocrine manner is present in these cells. Like its counterpart in the kidney, CYP27B1 occurring in monocytes and macrophages functions as a mixed function oxidase, with a similar affinity for 25(OH)D (Reichel et al. 1987). However, the regulation of CYP27B1 in these cells is completely different from that in the kidney. While the enzyme in the kidney is regulated by PTH and product feedback inhibition, the enzyme occurring in immune cells is not regulated by PTH or product inhibition but is strongly induced by an activation of the immune system (Adams et al. 1985; Reichel et al. 1987). Cytokines released by activation of the immune system such as TNF-a, IFN-y, interleukin (IL)-1, IL-2 and IL-15 are strong enhancers of the activity of CYP27B1 in monocytes and macrophages (Bouillon and Bikle 2019). 1,25(OH)2D resulting from the activation of CYP27B1 binds to VDR and stimulates the expression of genes involved in innate immunity including toll-like receptors 2 (TLR2) and -4 (TLR4) and CD14, a coreceptor of TLR4 (Oberg et al. 1993; Scherberich et al. 2005; Schauber et al. 2007). TLRrecognise pathogen-associated molecular patterns associated with infectious agents such as lipopolysaccharides (LPS) (Medzhitov 2007). TLR enhance vitamin D signalling by increasing the expression of CYP27B1 and VDR , constituting a positive feedback loop (Schauber et al. 2007). 1,25(OH)2D induces the formation of antimicrobial peptides such as the cathelicidin antimicrobial peptide (CAMP/LL37) and 6-defensin 2 (HBD2) (Bacchetta et al. 2014; Dimitrov and White 2016; Heulens et al. 2016; Lowry et al. 2020). Antimicrobial peptides exert a protective effect against a broad range of pathogens including fungi, bacteria, enveloped viruses and protozoa (Zasloff 2002). Besides their direct antimicrobial effect, they have an important immune modulatory activity, with either anti- or proinflammatory effects in different cells, depending on the inflammatory environment. They stimulate the recruitment of immune cells to the site of infection by interaction with specific receptors and are involved in tissue healing, revascularisation, cell proliferation and differentiation (Al-Rayahi and Sanyi 2015; Cristelo et al. 2021). Moreover, 1,25(OH)2D enhances the process of autophagy which leads to degradation of microbial proteins. Autophagy is induced by an inhibition of mTOR-pathway which represents an inhibitor of autophagy (Dimitrov et al. 2021). 1,25(OH)2D also stimulates the formation of superoxide anions in macrophages by an activation of NADPH- dependent phagocyte oxidase. The production of superoxide anions, leading to oxidative burst, is an important component in the degradation of bacteria (Sly et al. 2001). Recently, studies performed with macrophages have shown that 1,25(OH)2D stimulates the expression of the complement receptor immunoglobulin (CRIg), a unique complement receptor which plays a key role in the phagocytosis and clearance of bacteria (Small et al. 2021). On the other hand, 1,25(OH)2D acts in an anti-inflammatory manner as it increases the production of the anti-inflammatory IL-10 and decreases the release of pro- inflammatory mediators such as IL-16, IL-6, TNFa, receptor activator of nuclear factor kappa-B ligand (RANKL), and cyclo-oxygenase (COX)-2 (Martens et al. 2020; Saponaro et al. 2020). In mammary endothelial cells, it has been shown that 1,25(OH)2D attenuates stress of the endoplasmic reticulum (ER stress) and reduces the ER stress driven activation of NF-kB, supporting the view that vitamin D exerts anti-inflammatory effects (Wen et al. 2020).
In contrast to the innate immune system which is potentiated, the adaptive immune system is suppressed by 1,25(OH)2D. The adaptive immune response is based on B and T lymphocytes and their ability to produce cytokines and immunoglobulins in response to antigens presented to them (Bikle 2008). Mature dendritic cells play a major role for the presentation of antigens and the activation of T cell response. Mature dendritic cells are originating from monocytes, with immature dendritic cells as an intermediate state. Mature dendritic cells exert a high immune stimulating capacity. Their interaction with T cells leads to a T cell response. Depending on the formation of co-stimulating factors and the profile of interleukins released, a response of T helper cells (Th)1, Th2, Th17 or Th9 is induced. Dendritic cells can differentiate into immunogenic or tolerogenic cells, and hereby modulate T cell responses. Tolerogenic cells are characterised by a reduced expression of co-stimulatory molecules and a cytokine production favouring the induction of Th2 cells and regulatory T cells (formerly known as T suppressor cells) (Peelen et al. 2011; Martens et al. 2020). Regulatory T cells are immunosuppressive and generally suppress or downregulate induction and proliferation of effector T cells. They are also important in maintaining tolerance against self-antigens and prevent autoimmune diseases (Pulendran et al. 2010). Dendritic cells like Th and B lymphocytes express VDR , meaning that they are targets of 1,25(OH)2D. These cells also express CYP27B1, but only when they are activated. CYP27B1 in these cells, in difference to CYP27B1 in monocytes and macrophages, is inhibited by 1,25(OH)2D (Saponaro et al. 2020). Treatment with 1,25(OH)2D inhibits the differentiation of monocytes into immature dendritic cells as well as the maturation of immature dendritic cells into mature dendritic cells (Peelen et al. 2011). 1,25(OH)2D causes a decrease of the expression of the major histocompatibility complex (MHC) II, of CD80 and CD86, representing co-stimulatory molecules, as well as IL- 12 and IL-23. In contrast, inhibitory molecules such as immunoglobulin-like transcript (ILT)-3 and programmed death-ligand 1 (PD-L1) are upregulated. These changes enhance the induction of Th2 and regulatory T cells and suppress the activity of Th1 and Th17. 1,25(OH)2D therefore modulates dendritic cells towards a less mature and more tolerogenic phenotype with changes in morphology, cytokine production and surface markers (Martens et al. 2020; Saponaro et al. 2020). 1,25(OH)2D moreover decreases the production of the pro-inflammatory cytokines deriving from Th1 (IFN-y, IL-2), Th17 (IL-17, IL-21) and Th9 (IL-9) (Martens et al. 2020). In contrast, concentrations of IL-10 and the expression of forkhead box P3 (FoxP3), both of which are markers of regulatory T cells, are increased by 1,25(OH)2D (Martens et al. 2020). Incubation of B cells with 1,25(OH)2D inhibits their proliferation and retards their differentiation into plasma cells. In accordance with this, the release of immunoglobulins (Ig) M, G and A is suppressed by 1,25(OH)2D. Besides this direct effect, 1,25(OH)2D reduces the activation of T-lymphocytes by B-lymphocytes (Martens et al. 2020).
In sum, 1,25(OH)2D regulates both types of immune system. On the one hand, 1,25(OH)2D enhances the innate immunity and thus promotes the host's innate response to an invading pathogen. On the other hand, 1,25(OH)2D suppresses the adaptive immunity and thus prevents an overshooting response to the pathogen.
Findings of the effects of vitamin D and its metabolites on immune function in dairy cows and bovine cells
The number of studies dealing with the effect of vitamin D on the immune system in dairy cows is small in comparison to studies performed with human or murine immune cells. Most of the studies in cows had their focus on the effects of vitamin D or vitamin D metabolites [25(OH)D, 1,25(OH)2D] on the pathogenesis of mastitis. Investigation of the effects of vitamin D or its metabolites was mainly performed with bovine mammary epithelial cells (BMEC), immune cells isolated from blood of cows, or in mammary glands infected with pathogenic bacteria. In vivo studies also used somatic cells or immune cells from the mammary gland as samples for investigation of the effects of vitamin D or its metabolites on immunologic parameters.
It has been shown that BMEC as well as macrophages and neutrophils of cattle express both, CYP27B1 and VDR , and thus are enabled for an autocrine vitamin D signalling (Nelson et al. 2010a; Merriman et al. 2017, 2018; Yue et al. 2017; Poindexter et al. 2020). A study of Nelson et al. (2011) in which peripheral blood mononuclear cells (PBMC) of Holstein calves were sorted by a cell sorting system, showed that T cells, in contrast to B cells and macrophages have a negligible expression of CYP27B1; however, T cells had the highest expression of VDR among the three types of cells. The authors suggested that vitamin D signalling in T cells is induced by 1,25(OH)2D formed in monocytes or B cells in a paracrine manner (Nelson et al. 2011).
Several studies have shown that the activation of bovine immune cells (macrophages, neutrophils) by either infection with pathogenic bacteria (such as Streptococcus uberis) or by treatment with LPS causes an upregulation of CYP27B1 and VDR in these cells (Nelson et al. 2010a, 2010b, 2011; Merriman et al. 2018). These findings show that, like in human or murine immune cells, activation of immune cells of cattle leads to a stimulation of vitamin D signalling by an enhanced formation of 1,25(OH)2D and an upregulation of VDR . Based on this observation, several studies investigated the hypothesis that administration of vitamin D or its hydroxylated metabolites is able to improve immune function in bovine cells.
Lippolis et al. (2011) studied the effect of the injection of 100 ug 25(OH)D3 into a quarter of the mammary gland of dairy cows infected with Streptococcus uberis. In this study, infection of the mammary gland with Streptococcus uberis led to increases in bacteria count and number of somatic cells in the milk, body temperature and concentration of bovine serum albumin in the milk, and to a reduction of feed intake. Administration of 25(OH)D3 into the mammary gland improved all these parameters either in a significant manner, or there were at least tendencies towards an improvement. Plasma concentration of 25(OH)D of the cows in this experiment was between 50 and 60 ng/ml, indicating that the cows had a sufficient vitamin D status. This indicates that 25(OH)D3 injected exerted local effects in the mammary gland affected. This study was the first one which indicated that vitamin D or its metabolites could exert beneficial effects in the prevention or therapy of mastitis in dairy cows. In a similar experimental design, Merriman et al. (2017) injected 10 ug 1,25(OH)2D3 into the mammary gland of cows free of mastitis or cows with subclinical mastitis. They determined expression of several genes in milk somatic cells including defensins (DEFB3, DEFB4, DEFB5, DEFB7, DEFB10), cytokines (C-C motif chemokine ligand 5 (CCL5), IL-1B, IL-8) and inducible nitric oxide synthase (iNOS). In healthy cows, the only change observed by 1,25(OH)2D3- injection was an upregulation of iNOS, an enzyme which produces nitric oxide. In cows with subclinical mastitis, 1,25(OH)2D3 injection caused an increase of the expression of DEFB4, a member of the family of 6-defensins, and iNOS. Nitric oxide, which is produced by iNOS, and 6-defensins are known to play an important role in the protection against bacterial pathogens (Bogdan 2001; Solanki et al. 2021). The authors of this study concluded that vitamin D metabolites could be useful in mastitis treatment by improving the antibacterial competence of mammary gland cells. In the study of Merriman et al. (2018), mammary glands of cows with an adequate vitamin D status [43 ng 25(OH)D/ml plasma] were injected with LPS and subsequently with 100 ug 25(OH)D3 in order to investigate the hypothesis that 25(OH)D administration ameliorates the inflammation induced by LPS. As expected, injection of LPS caused a strong immune response (increase of the expression of defensins, CCL5, iNOS, Il-1B, IL-8) in somatic cells, macrophages and neutrophils in the milk. However, treatment with 25(OH)D3 had no effect on the inflammation process induced by LPS. The authors suggested that the lack of effect of 25(OH)D3 on immune response in the mammary gland is due to the fact that the immune reaction induced by LPS occurs much faster than an immune reaction induced by infection with bacteria.
Besides the studies in which cows were treated with 25(OH)D3 or 1,25(OH)2D3 by administration into the mammary gland, there are very few studies available so far which investigated the effect of oral supplementation of vitamin D or vitamin D metabolites on the immune system of the mammary gland in dairy cows. Poindexter et al. (2020) supplemented four groups of lactating dairy cows with either 1 or 3 mg of vitamin D3 or 1 or 3 mg 25(OH)D3 per day for a period of 28 d. At day 21, cows supplemented with 1 mg vitamin D3 or 3 mg 25(OH)D3 were additionally subjected to an intramammary challenge with Streptococcus uberis. Cows supplemented with 1 or 3 mg of vitamin D3 had 25(OH)D-concentrations in plasma within a normal range, such as those observed in practical dairy herds [63 and 68 ng 25(OH)D/ml plasma, resp.]. In contrast cows supplemented with 1 or 3 mg of 25(OH)D3 had extremely high concentrations of 25 (OH)D in plasma which are in clear excess of values observed in practical dairy herds (140 and 230 ng/ml plasma, resp.). Milk yield and dry matter intake were not different between the four groups of cows. In this study, the expression of several genes involved in immune response in milk somatic cells before and after immunologic challenge was determined. Before challenge, the only genes influenced by treatment were IL-1B and iNOS; the expression levels of those genes were increased in the groups treated with 25(OH)D3 in comparison to the groups treated with vitamin D3. There were positive correlations between the plasma concentration of 25(OH)D and of the expression of some defensins (DEFB7, DEFB10), IL-1B and iNOS. The expression of these genes was moreover also positively correlated with the expression of CYP27B1 and VDR , indicating that the upregulation of these genes involved in immune response changes were induced by 1,25(OH)2D mediated vitamin D signalling. After challenge, the group supplemented with a daily dose of 3 mg 25(OH)D3 showed less severe symptoms of mastitis whereas milk yield, feed intake, rectal temperature, somatic cell count, bacterial count and concentration of bovine serum albumin in the milk were not different from the group treated with 1 mg vitamin D3. The expression levels of the adhesion protein CD11b on milk neutrophils and the expression of iNOS in milk somatic cells were increased in the group supplemented with 3 mg 25(OH)D3 in comparison to the group supplemented with 1 mg vitamin D3. The increased expression of CD11b in somatic cells is an indication of an increased leucocyte recruitment to the mammary gland upon bacterial challenge. Overall, the findings of this study support the hypothesis that vitamin D signalling could play role in the prevention of mastitis of dairy cows. In a more recent study of the same group (Vieira-Neto et al. 2021), cows were supplemented with either vitamin D3 (1 or 3 mg/d) or 25(OH)D3 (1 or 3 mg/d) from day 250 of gestation until calving. As expected supplementation with vitamin D3 caused only a moderate increase of plasma concentration of 25(OH)D (about 60-75 ng/ml) whereas treatment with 25(OH)D3 yielded much higher of 25(OH)D concentrations in plasma during the treatment period [85-100 ng/ml in the group treated with 1 mg 25(OH)D3 per day, 150-210 ng/ml in the group treated with 3 mg 25(OH)D3 per day]. Treatment with both doses of 25(OH)D3 increased mRNA concentrations of various genes in leucocytes which are involved in the immune defence including several genes related to cell adhesion and migration, pathogen recognition receptor, cell signalling, cytokine signalling and antimicrobial mechanism in comparison to treatment with vitamin D3 prepartum and postpartum. However, although there were significant effects on the expression of several genes involved in the immune system, there were no differences in the incidence of diseases (retained placenta, metritis, mastitis) between the four groups during the first 60 d postpartum. This means that despite of the observed changes of the concentrations of 25(OH)D in plasma and the changes of mRNA concentrations of genes involved in immune function in leucocytes, the treatment with 25(OH)D3 was unable to affect the risk of clinical diseases in the early lactation period. Nevertheless, this study supports the view that vitamin D is involved in immune function of dairy cows. Moreover, the study confirms that effects on immune function are induced only by supraphysiological plasma concentration of 25(OH)D which serves as a substrate for the local production of 1,25(OH)2D in leucocytes. It is likely that the upregulation of the expression of genes involved in immune defence by treatment with 25(OH)D3 observed in this study was induced by an enhancement of vitamin D signalling in leucocytes. In the study of Yue et al. (2018) healthy dairy cows were deprived of vitamin D for 15 weeks by omitting vitamin D supplements from their feed and housing them without access to sunlight. Deprivation of vitamin D yielded plasma 25(OH)D concentrations lower than 10 ng/ml. Beginning at day 219 of lactation, the cows were then repleted with vitamin D by either supplementation of vitamins D2 or D3 or by exposition to sunlight for 29 d. The repletion treatment led to 25(OH)D plasma concentrations of about 30 to 45 ng/ml. However, none of the immune parameters determined in this study (leucocyte populations, somatic cell count, immunoglobulin concentrations in plasma and milk, antigen-stimulated cytokine productions) did change in response to vitamin D repletion or vitamin D source. It is likely that the lack of effect of vitamin D repletion on the immune function was mainly due to the fact that the cows in the late lactation were in a healthy condition and were not subjected to inflammatory stress.
Besides the investigations performed in cows in vivo, there are some studies dealing with the effects of vitamin D or its metabolites on the immune response in vitro in either BMEC or various types of immune cells.
Nelson et al. (2010b) have shown that treatment of LPS stimulated bovine monocytes with 1,25(OH)2D causes a strong, dose-dependent, increase of the expression of iNOS und RANTES (regulated upon activation, normal T cell expressed and secreted) and the production of nitric oxide. Nitric oxide, as mentioned above, is an important component of the antimicrobial response, while RANTES is a chemo-attractant for T-helper cells and monocytes and thus is important for the recruitment of other immune cells to the site of inflammation (Levy 2009). An upregulation of iNOS by 1,25(OH)2D was also shown in primary bovine dermal fibroblasts treated with LPS and in primary bovine salivary gland fibroblasts treated with LPS (Boylan et al. 2020; O’Brien et al. 2021). Merriman et al. (2015) observed that the incubation of bovine monocyte cultures with 1,25(OH)2 D causes a strong (5-10-fold) upregulation of various members of the bovine 6- defensin family and of iNOS, both in the absence or presence of LPS. The authors of that study suggested that the upregulation of 6-defensins by 1,25(OH)2D could be due to potentiating of NF-kB-signalling. Nelson et al. (2011) investigated the effect of vitamin D metabolites in PBMC of calves subjected to a purified protein derivative of Mycobacterium bovis. In this study, treatment with 25(OH)D or 1,25(OH)2D caused an upregulation of iNOS and RANTES and a downregulation of IFN-y and IL-17. Sorting of the PBMCs into monocytes, B cell and T cell fractions has shown that the upregulation of iNOS and RANTES was mainly induced in monocytes, and to a less extent in B cells, while the downregulation of IFN-y and IL-17 was mainly induced in T cells. A reduced production of IFN-y in bovine mononuclear leukocytes by treatment with 1,25(OH)2 D has also been reported in earlier studies (Ametaj et al. 1996; Waters et al. 2001; Nonnecke et al. 2003).
Two studies investigated the effect of vitamin D on the internalisation of Staphylococcus aureus into BMEC. In the study of Tellez-Perez et al. (2012) incubation with vitamin D lowered the internalisation of Staphylococcus aureus into BMEC. The authors of this study suggested that this effect was induced by an autocrine action of 1,25(OH)2D, as BMEC express both a 25-hydroxylase (CYP2R1) and a 1-hydroxylase (CYP27B1) and thus are capable of producing 1,25(OH)2D from vitamin D. In agreement with that study, the study of Yue et al. (2017) observed that treatment with either 25(OH)D2 or 25(OH)D3 reduces the invasion of Staphylococcus aureus into BMEC. However, the inhibitory effect of 25(OH)D3 on invasion was greater than that of 25(OH)D2. Mechanisms by which vitamin D or vitamin D metabolites inhibit the invasion of Staphylococcus aureus into BMEC were not elucidated in these studies.
Effects of Vitamin D on the immune function of dairy cows: conclusion
The number of studies investigating the effects of vitamin D or vitamin D metabolites on immune function in dairy cows and bovine cells is very limited. However, the studies available so far give clear evidence that the expression of CYP27B1 and VDR in bovine immune cells is stimulated by immune activators such as LPS or bacterial products (Nelson et al. 2010a, 2010b, 2011; Merriman et al. 2018). This finding, which agrees with observations in human and murine immune cells, shows that an activation of bovine immune cells leads to a stimulation of vitamin D signalling. Yet now, there are few studies available dealing with effects of vitamin D or vitamin D metabolites on immune function in dairy cows. There is one study showing that an administration of 25(OH)D3 into an infected mammary gland affects the pathogenesis of mastitis in a beneficial manner (Lippolis et al. 2011). Several studies in cows and bovine cells consistently show that treatment with 25(OH)D3 (in cows) or 1,25(OH)2D3 (in bovine cells) increases the expression of iNOS and 6-defensins (Nelson et al. 2010b, 2011; Merriman et al. 2015, 2017; Boylan et al. 2020; Poindexter et al. 2020; O’Brien et al. 2021). In two studies, treatment with 1,25(OH)2D3 additionally stimulated the expression of RANTES (Nelson et al. 2010b, 2011). These findings agree with observations made in human or murine immune cells, indicating that treatment with vitamin D metabolites enhances the innate immune defence. Therefore, beneficial effects of 25(OH)D3 on the pathogenesis of mastitis might be due to an improvement of the defence against bacteria by the production of antimicrobial peptides and nitric oxide. A model of the effects of vitamin D on immune cells (macrophages, monocytes) of dairy cows, including recent findings, is presented in Figure 2.
Figure 2. Model of the effects of vitamin D on bovine immune
cells (macrophages, monocytes) and mammary epithelial cells. Pathogen-associated molecular patterns (PAMP) like lipopolysaccharides (LPS) bind to their receptors like toll-like receptors (TLR), and thereby initiate an immune response in the cell. The initiation of the immune response leads to an upregulation of CYP27B1 and vitamin D receptor ( VDR ). Activation of CPY27B1 causes an increased production of 1,25-dihydroxyvitamin D [1,25(OH)2D] from its precursor 25-hydroxyvitamin D [25(OH)D] which is taken up from the blood. 1,25(OH)2D then binds to the VDR , which after complexing with retinoid X receptor (RXR) and other cofactors, binds to vitamin D response element ( VDR E) and induces the transcription of “regulated and normal T cells expressed and secreted“ (RANTES), “inducible nitric oxide synthase“ (iNOS) and B- defensins, representing genes known to improve the innate immune response. For more details and references, see text.
The finding of a correlation between plasma 25(OH)D concentration and the expression of defensins and iNOS strongly suggest that the upregulation of the genes is induced by 25(OH)D taken up into immune cells or epithelial cells of the mammary gland (Poindexter et al. 2020). Beneficial effects of 25(OH)D were observed in cows which - according to their plasma 25(OH)D concentrations - had already a sufficient vitamin D status prior to treatment. This finding indicates that beneficial effects of 25(OH)D might be induced particularly at supraphysiological concentrations. Such high concentrations are not reached by oral supply of vitamin D, but only by supply of 25(OH)D. In the study of Poindexter et al. (2020), cows supplemented daily with 3 mg 25(OH)D3 showed a more favourable pathogenesis of mastitis than cows supplemented daily with 1 mg vitamin D3. Investigations in which cows were injected with 25(OH)D3 into the mammary gland suggest that high local concentrations of 25(OH)D in a tissue could exert beneficial effects on the immune system of this tissue. It is likely that high local concentrations of 25(OH)D mediate their effects on the immune function by an enhancement of the local production of 1,25(OH)2D which in turn activates the expression of protective genes such as iNOS or 6-defensins. However, according to the study of Vieira- Neto et al. (2021), an improvement of the immune defence induced by high levels of 25 (OH)D does not necessarily prevent against the development of infectious diseases during early lactation. The concentration of 25(OH)D required for optimising the innate immune defence remains unknown due to the lack of a sufficient number of studies, including dose-response trials. Another finding in bovine mononuclear leucocytes wasthat treatment with 1,25(OH)D2 causes a reduction of the production of IFN-y (Ametaj et al. 1996; Waters et al. 2001; Nonnecke et al. 2003; Nelson et al. 2011). This finding agrees with studies in human and murine lymphocytes showing that treatment with 1,25(OH)2D3 leads to a suppression of the adaptive immune system (Peelen et al. 2011). However, studies in dairy cows dealing with effects of vitamin D or its metabolites on the adaptive immune system are lacking so far.
Conclusion and future directions
The function of vitamin D in calcium homoeostasis in dairy cows, such as in other vertebrates, is known for many years. Studies in the last decade yielded some indication that vitamin D is also important for the function of immune cells in dairy cows, and thus could be useful in the prevention and therapy of infectious diseases such as mastitis. However, data available in this respect are very limited due to the small number of trials in dairy cows performed so far. Therefore, further investigations of the effects of vitamin D or its metabolite 25(OH)D on the immune function in dairy cows are required. In particular, there is a need for dose-response studies in order to determine the dose of vitamin D or 25(OH)D which is required to support immune function. Plasma concentration of 25(OH)D is regarded as an indicator of the vitamin D status in dairy cows as in other species. 25(OH)D is formed from vitamin D by 25-hydroxylases in the liver. Studies in humans and mice have shown that 25-hydroxylation of vitamin D and resulting plasma concentrations of 25(OH)D are influenced by various factors including BW, age or diet. In dairy cows, there are no studies so far dealing with the regulation of 25- hydroxylation in the liver. However, the observation of a relationship between the existence of SNP in CYP27R1, the enzyme which catalyses the formation of 25(OH)D from vitamin D in the liver, and the incidence of clinical mastitis already indicates that 25-hydroxylation of vitamin D in the liver might be relevant for immune function. Recent studies moreover strongly suggest that non-classical vitamin function which are based on an autocrine vitamin D signalling are more depending on the concentration of free 25(OH)D than on the concentration of total 25(OH)D in plasma, according to the “free hormone hypothesis”. Concentrations of free 25(OH)D in the plasma of dairy cows have not yet been determined. However, the concentration of free 25(OH)D depends on the concentration and the binding affinity of DBP . The binding affinity to 25(OH)D could be influenced by polymorphisms in the DBP gene. There is at least one study in dairy cows which identified DBP as a strong candidate gene associated with incidence of clinical mastitis, which indicated that SNP in DBP could have an impact on the immune system via an influence on the concentration of free 25(OH)D. Therefore, DBP could be an interesting target for further investigations in dairy cows. Recent studies have shown that DBP has a lower binding affinity for 25(OH)D2 than for 25(OH)D3, meaning that supplementation of vitamin D2 yields higher concentrations of free 25(OH)D than supplementation of vitamin D3. Thus, vitamin D2 could enhance vitamin D function in cells with autocrine vitamin D signalling (such as immune cells), and thus could be an interesting alternative to vitamin D3 for supplementation in dairy cows. The functions of vitamin D are mediated by VDR which also could be an interesting target for further investigation. In humans, it has been shown that vitamin D signalling is strongly influenced by polymorphisms in the VDR gene, associated with an increased incidence of various diseases. Therefore, search for polymorphisms in the VDR gene with respect to a possible relationship to functions of vitamin D would be of interest. Overall, it is clear that vitamin D is highly relevant for the immune function in dairy cows. However, many open questions need to be answered, in order to enable an optimised usage of vitamin D in dairy cows for the improvement of the immune function.
References
Aatsinki S-M, Elkhwanky M-S, Kummu O, Karpale M, Buler M, Viitala P, Rinne V, Mutikainen M, Tavi P, Franko A, et al. 2019. Fasting-induced transcription factors repress vitamin D bioactivation, a mechanism for vitamin D deficiency in diabetes. Diabetes. 68:918-931.
Abboud M, Puglisi DA, Davies BN, Rybchyn M, Whitehead NP, Brock KE, Cole L, Gordon- Thomson C, Fraser DR, Mason RS. 2013. Evidence for a specific uptake and retention mechanism for 25-hydroxyvitamin D (25OHD) in skeletal muscle cells. Endocrinology. 154:3022-3030.
Abboud M, Rybchyn MS, Liu J, Gordon-Thomson C, Brennan-Speranza TC, Cole L, Greenfield H, Fraser DR, Mason RS. 2017. The effect of parathyroid hormone on the uptake and retention of 25-hydroxyvitamin D in skeletal muscle cells. J Steroid Biochem Mol Biol. 173:173-179.
Abdel-Shafy H, Bortfeldt RH, Reissmann M, Brockmann GA. 2018. Validating genome-wide associated signals for clinical mastitis in German Holstein cattle. Anim Genet. 49:82-85.
Adams JS, Singer FR, Gacad MA, Sharma OP, Hayes MJ, Vouros P, Holick MF. 1985. Isolation and structural identification of 1,25-dihydroxyvitamin D3 produced by cultured alveolar macrophages in sarcoidosis. J Clin Endocrinol Metab. 60:960-966.
Aiba I, Yamasaki T, Shinki T, Izumia S, Yamamoto K, Yamada S, Terato H, Ide H, Ohyama Y. 2006. Characterization of rat and human CYP2J enzymes as Vitamin D25-hydroxylases. Steroids. 71:849-856.
Al-Rayahi IAM, Sanyi RHH. 2015. The overlapping roles of antimicrobial peptides and complement in recruitment and activation of tumor-associated inflammatory cells. Fron Immunol. 6:2.
Ametaj BN, Beitz DC, Reinhardt TA, Nonnecke BJ. 1996. 1,25-Dihydroxyvitamin D3 inhibits secretion of interferon-y by mitogen- and antigen-stimulated bovine mononuclear leukocytes. Veter Immunol Immunopathol. 52:77-90.
Anderson PH, Atkins GJ. 2008. The skeleton as an intracrine organ for vitamin D metabolism. Mol Aspects Med. 29:397-406.
Arnaud J, Constans J. 1993. Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein ( DBP ). Hum Genet. 92:183-188.
Bacchetta J, Chun RF, Gales B, Zaritsky JJ, Leroy S, Wesseling-Perry K, Boregard N, Rastogi A, Salusky IB, Hewison M . 2014. Antibacterial responses by peritoneal macrophages are enhanced following vitamin D supplementation. PLoS One. 9:e116530.
Baldock PA, Thomas GP, Hodge JM, Baker SU, Dressel U, O’Loughlin PD, Nicholson GC, Briffa KH, Eisman JA, Gardiner EM. 2006. Vitamin D action and regulation of bone remodeling: suppression of osteoclastogenesis by the mature osteoblast. J Bone Miner Res. 21:1618-1626.
Baur AC, Brandsch C, Steinmetz B, Schutkowski A, Wensch-Dorendorf M, Stangl GI. 2020. Differential effects of vitamin D3 vs vitamin D2 on cellular uptake, tissue distribution and activation of vitamin D in mice and cells. J Steroid Biochem Mol Biol. 204:105768.
Bikle DD. 2008. Vitamin D and the immune system: role in protection against bacterial infection. Curr Opin Nephrol Hypertens. 17:348-352.
Bikle D. 2009. Nonclassic actions of vitamin D. J Clin Endocrinol Metab. 94:26-34.
Bikle DD. 2020. The free hormone hypothesis: when, why, and how to measure the free hormone levels to assess vitamin D, thyroid, sex hormone, and cortisol status. JBMR Plus. 5:e10418.
Bikle D, Bouillon R, Thadhani R, Schoenmakers I. 2017a. Vitamin D metabolites in captivity? Should we measure free or total 25(OH)D to assess vitamin D status? Steroid Biochem Mol Biol. 173:105-116.
Bikle DD, Halloran BP, Gee E, Ryzen E, Haddad JG. 1986. Free 25-hydroxyvitamin D levels are normal in subjects with liver disease and reduced total 25-hydroxyvitamin D levels. J Clin Invest. 78:748-752.
Bikle DD, Malmstroem S, Schwartz J. 2017b. Current controversies: are free vitamin metabolite levels a more accurate assessment of vitamin D Status than total levels? Endocrinol Metab Clin North Am. 46:901-918.
Bikle DD, Schwartz J. 2019. Vitamin D binding protein, total and free vitamin D levels in different physiological and pathophysiological conditions. Front Endocrinol (Lausanne). 10:317.
Bogdan C. 2001. Nitric oxide and the immune response. Nat Immunol. 2:907-916.
Boros S, Bindels RJ, Hoenderop JG. 2009. Active Ca(2+) reabsorption in the connecting tubule. Pflugers Arch. 458:99-109.
Bouillon R, Bikle D. 2019. Vitamin D metabolism revised: fall of dogmas. J Bone Miner Res. 34:1985-1992.
Bouillon R, Schuit F, Antonio L, Rastinejad F. 2020. Vitamin D binding protein: a historic overview. Front Endocrinol (Lausanne). 10:910.
Bouillon R, Xiang DZ, Convents R, Van Baelen H. 1992. Polyunsaturated fatty acids decrease the apparent affinity of vitamin D metabolites for human vitamin D-binding protein. J Steroid Biochem Mol Biol. 42:855-861.
Boyce BF, Xing L. 2008. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch Biochem Biophys. 473:139-146.
Boylan M, O’Brien MB, Beynon C, Meade KG. 2020. 1,25(OH)D vitamin D promotes NOS2 expression in response to bacterial and viral PAMPs in primary bovine salivary gland fibroblasts. Vet Res Commun. 44:83-88.
Bradford BJ, Yuan K, Farney JK, Mamedova LK, Carpenter AJ. 2015. Invited review: inflammation during the transition to lactation: new adventures with an old flame. J Dairy Sci. 98:6631-6650.
Carlsson M, Nilsson I, Brudin L, Von SP, Wanby P. 2017. Erythrocyte fatty acid composition does not influence levels of free, bioavailable, and total 25-hydroxy vitamin D. Scand J Clin Lab Invest. 77:45-52.
Christakos S, Dhawan P, Porta A, Mady LJ, Seth T. 2011. Vitamin D and intestinal calcium absorption. Mol Cell Endocrinol. 347:25-29.
Chun RF, Hernandez I, Pereira R, Swinkles L, Huijs T, Zhou R, Liu NQ, Shieh A, Guemes M, Mallya SM, et al. 2016. Differential responses to vitamin D2 and vitamin D3 are associated with variations in free 25-hydroxyvitamin D. Endocrinology. 157:3420-3430.
Chun RF, Shieh A, Gottlieb C, Yacoubian V, Wang J, Hewison M, Adams JS. 2019. Vitamin D binding protein and the biological activity of vitamin D. Front Endocrinol (Lausanne). 10:718.
Contreras GA, O’Boyle NJ, Herdt TH, Sordillo LM. 2010. Lipomobilization in periparturient dairy cows influences the composition of plasma nonesterified fatty acids and leukocyte phospholipid fatty acids. J Dairy Sci. 93:2508-2516.
Cristelo C, Machado A, Sarmento B, Gama FM. 2021. The roles of vitamin D and cathlecidin in type 1 diabetes susceptibility. Endocr Connect. 10:R1-R12.
Deiner C, Reiche M, Lassner D, Grienitz D, Twardziok S, Moesch A, Wenning P, Martens H. 2012. Allelic variations in coding regions of the vitamin D receptor gene in dairy cows and potential susceptibility to periparturient hypocalcaemia. J Dairy Res. 79:423-428.
Dimitrov V, Barbier C, Ismailova A, Wang Y, Dmowski K, Salehi-Tabar R, Memari B, Groulx- Boivin E, White JH. 2021. Vitamin D-regulated gene expression profiles: species-specificity and cell-specific effects on metabolism and immunity. Endocrinology. 162:bqaa218.
Dimitrov V, White JH. 2016. Species-specific regulation of innate immunity by vitamin D signaling. J Steroid Biochem Mol Biol. 164:246-253.
Dittmer KE, Thompson KG. 2011. Vitamin D metabolism and rickets in domestic animals: a review. Vet Pathol. 48:389-407.
Duchow EG, Cooke NE, Seeman J, Plum LA, DeLuca HF. 2019. Vitamin D binding protein is required to utilize skin-generated vitamin D. Proc Natl Acad Sci U S A. 116:24527-24532.
Dueland S, Bouillon R, Van Baelen H, Pedersen JI, Helgerud P, Drevon CA. 1985. Binding protein for vitamin D and its metabolites in rat mesenteric lymph. Am J Physiol. 249:E1-5.
Dueland S, Pedersen JI, Helgerud P, Drevon CA. 1983. Absorption, distribution, and transport of vitamin D3 and 25-hydroxyvitamin D3 in the rat. Am J Physiol. 245:E463-467.
Ekwaru JP, Zwicker JD, Holick MF, Giovannucci E, Veugelers PJ. 2014. The importance of body weight for the dose response relationship of oral vitamin D supplementation and serum 25-hydroxyvitamin D in healthy volunteers. PLoS One. 9:e111265.
Garbossa SG, Folli F. 2017. Vitamin D, sub-inflammation and insulin resistance. A window on a potential role for the interaction between bone and glucose metabolism. Rev Endocr Metab Disord. 18:243-258
Gessner DK, Koch C, Romberg FJ, Winkler A, Dusel G, Herzog E, Most E, Eder K. 2015. The effect of grape seed and grape marc meal extract on milk performance and the expression of genes of endoplasmic reticulum stress and inflammation in the liver of dairy cows in early lactation. J Dairy Sci. 98:8856-8868.
Gessner DK, Schlegel G, Keller J, Schwarz FJ, Ringseis R, Eder K. 2013. Expression of target genes of nuclear factor E2-related factor 2 in the liver of dairy cows in the transition period and at different stages of lactation. J Dairy Sci. 96:1038-1043.
Gnagnarella P, Raimondi S, Aristarco V, Johansson H, Bellerba F, Corso F, De Angelis SP, Belloni P, Caini S, Gandini S. 2021. Ethnicity as modifier of risk for vitamin D receptors polymorphisms: comprehensive meta-analysis of all cancer sites. Crit Rev Oncol Hematol. 158:103202.
Golder HM, McGrath J, Lean IJ. 2021. Effect of 25-hydroxyvitamin D3 during prepartum transition and lactation on production, reproduction, and health of lactating dairy cows. J Dairy Sci. 104:5345-5374.
Guillot X, Semerano L, Saidenberg-Kermanach N, Falgarone G, Boissier M-C. 2010. Vitamin D and inflammation. Joint Bone Spine. 77:552-557.
Heaney RP, Recker RR, Grote J, Horst RL, Armas LA. 2011. Vitamin D3 is more potent than vitamin D2 in humans. Clin Endocrinol Metab. 96:E447-52.
Henderson CM, Fink SL, Bassyouni H, Argiropoulos B, Brown L, Laha TJ, Jackson KJ, Lewkonia R, Ferreira P, Hoofnagle AN, et al. 2019. Vitamin D-binding protein deficiency and homozygous deletion of the GC gene. N Engl J Med. 380:1150-1157.
Heulens N, Korf H, Mathyssen C, Everaerts S, De Smidt E, Dooms C, Yserbyt J, Gysemans C, Gayan-Ramirez G, Mathieu C, et al. 2016. 1,25-dihydroxyvitamin D modulates antibacterial and inflammatory response in human cigarette smoke-exposed macrophages. PLoS One. 11: e0160482.
Hidiroglou M, Proulx JG, Roubos D. 1979. 25-hydroxyvitamin D in plasma of cattle. J Dairy Sci. 62:1076-1080.
Hodnik JJ, Jezek J, Staric J. 2020. A review of vitamin D and its importance to the health of dairy cattle. J Dairy Res. 87:84-87.
Holcombe SJ, Wisnieski L, Gandy J, Norby B, Sordillo LM. 2018. Reduced serum vitamin D concentrations in healthy early-lactation dairy cattle. J Dairy Sci. 101:1488-1494.
Hollis BW. 1984. Comparison of equilibrium and disequilibrium assay conditions for ergocalci- ferol, cholecalciferol and their major metabolites. J Steroid Biochem. 21:81-86.
Horst RL, Goff JP, Reinhardt TA. 1994. Calcium and vitamin D metabolism in the dairy cow. J Dairy Sci. 77:1936-1951.
Horst RL, Reinhardt TA. 1983. Vitamin D metabolism in ruminants and its relevance to the periparturient cow. J Dairy Sci. 66:661-678.
Huhtakangas JA, Olivera CJ, Bishop JE, Zanello LP, Norman AW. 2004. The vitamin D receptor is present in caveolae-enriched plasma membranes and binds 1 alpha,25(OH)2-vitamin D3 in vivo and in vitro. Mol Endocrinol. 18:2660-2671.
Hym0ller L, Jensen SK. 2010a. Stability in the rumen and effect on plasma status of single oral doses of vitamin D and vitamin E in high-yielding dairy cows. J Dairy Sci. 93:5748-5757.
Hym0ller L, Jensen SK. 2010b. Vitamin D3 synthesis in the entire skin surface of dairy cows despite hair coverage. J Dairy Sci. 93:2025-2029.
Hym0ller L, Jensen SK. 2012. 25-Hydroxycholecalciferol status in plasma is linearly correlated to daily summer pasture time in cattle at 56°N. Br J Nutr. 108:666-671.
Hym0ller L, Jensen SK, Kaas P, Jakobsen J. 2017. Physiological limit of the daily endogenous cholecalciferol synthesis from UV light in cattle. J Anim Physiol Anim Nutr (Berl). 101:215-221.
Hym0ller L, Jensen SK, Lindqvist H, Johansson B, Nielsen MO, Nadeau E. 2009. Supplementing dairy steers and organically managed dairy cows with synthetic vitamin D3 is unnecessary at pasture during exposure to summer sunlight. J Dairy Res. 76:372-378.
Ismailova A, White JH. 2021. Vitamin D, infections and immunity. Rev Endocr Metab Disord. 29:1-13.
Jacquillet G, Unwin R. 2019. Physiological regulation of phosphate by vitamin D, parathyroid hormone (PTH) and phosphate (Pi). Pflügers Arch. 471:83-98.
Jäpelt RB, Jakobsen J. 2013. Vitamin D in plants: a review of occurrence, analysis, and biosynthesis. Front Plant Sci. 4:136.
Jones KS, Assar S, Harnpanich D, Bouillon R, Lambrechts D, Prentice A, Schoenmakers I. 2014b. 25(OH)D2 half-life is shorter than 25(OH)D3 half-life and is influenced by DBP concentration and genotype. J Clin Endocrinol Metab. 99:3373-3381.
Jones G, Byrnes B, Palma F, Segev D, Mazur Y. 1980. Displacement potency of vitamin D2 analogs in competitive protein-binding assays for 25-hydroxyvitamin D3, 24,25-dihydroxyvitamin D3, and dihydroxyvitamin D3. J Clin Endocrinol Metab. 50:773-775.
Jones G, Prosser DE, Kaufmann M. 2014a. Cytochrome P450-mediated metabolism of vitamin D. J Lipid Res. 55:13-31.
Kiela PR, Ghishan FK. 2016. Physiology of intestinal absorption and secretion. Best Pract Res Clin Gastroenterol. 30:145-159.
Koivisto O, Hanel A, Carlberg C. 2020. Key vitamin D target genes with functions in the immune system. Nutrients. 12:1140.
Kutuzova GD, Deluca HF. 2004. Gene expression profiles in rat intestine identify pathways for
dihydroxyvitamin D3 stimulated calcium absorption and clarify its immunomodulatory properties. Arch Biochem Biophys. 432:152-166.
Lehmann U, Hirche F, Stangl GI, Hinz K, Westphal S, Dierkes J. 2013. Bioavailability of vitamin D2 and D3 in healthy volunteers, a randomized placebo-controlled trial. J Clin Endocrinol Metab. 98:4339-4345.
Levy JA. 2009. The unexpected pleiotropic activities of RANTES. J Immunol. 182:3945-3946.
Li H-M, Liu Y, Zhang R-J, Ding J-Y, Shen C-L. 2021. Vitamin D receptor gene polymorphisms and osteoarthritis: a meta-analysis. Rheumatology (Oxford). 60:538-548.
Lippolis JD, Reinhardt TA, Sacco RA, Nonnecke BJ, Nelson CD. 2011. Treatment of an intramammary bacterial infection with 25-hydroxyvitamin D3. PLoS One. 6:e25479.
Littledike eT, Horst RL. 1982. Vitamin D3 toxicity in dairy cows. J Dairy Sci. 65:749-759.
Liu J, Greenfield H, Strobel N, Fraser DR. 2013. The influence of latitude on the concentration of vitamin D3 and 25-hydroxy-vitamin D3 in Australian red meat. Food Chem. 140:432-435.
Liu L, Shen T, Yang W, Yu H, Gao S, Huang B, Xu C. 2020. Ketotic cows display a different serum nonesterified fatty acid composition. J Dairy Res. 87:52-55.
Louw JA, Werbeck A, Louw ME, Kotze TJ, Cooper R, Labadarios D. 1992. Blood vitamin concentrations during the acute-phase response. Crit Care Med. 20:934-941.
Lowry BM, Guo C, Zhang Y, Fantacone ML, Logan IE, Campbell Y, Zhang W, Le M, Indra AK, Ganguli-Indra G, et al. 2020. A mouse model for vitamin D-induced human cathelicidin antimicrobial peptide gene expression. J Steroid Biochem Mol Biol. 198:105552.
Makris K, Sempos C, Cavalier E. 2020. The measurement of vitamin D metabolites: part I-metabolism of vitamin D and the measurement of 25-hydroxyvitamin D. Hormones (Athens). 19:81-96.
Mangin M, Sinha R, Fincher K. 2014. Inflammation and vitamin D: the infection connection. Inflamm Res. 63:803-819.
Marcinowska-Suchowierska E, Kupisz-Urbanska M, Lukaszkiewicz J, Pludowski P, Jones G. 2018. Vitamin D toxicity-a clinical perspective. Front Endocrinol (Lausanne). 9:550.
Margier M, Collet X, Le May C, Desmarchelier C, André F, Lebrun C, Defoort C, Bluteau A, Borel P, Lespine A, et al. 2019. ABCB1 (P-glycoprotein) regulates vitamin D absorption and contributes to its transintestinal efflux. FASEB J. 33:2084-2094.
Martens PJ, Gysemans C, Verstuyf A, Mathieu AC. 2020. Vitamin D’s effect on immune function. Nutrients. 12:1248.
Medzhitov R. 2007. Recognition of microorganisms and activation of the immune response. Nature. 449:819-826.
Mello JRB. 2003. Calcinosis-calcinogenic plants. Toxicon. 41:1-12.
Merriman KE, Kweh MF, Powell JL, Lippolis JD, Nelson CD. 2015. Multiple ß-defensin genes are upregulated by the vitamin D pathway in cattle. J Steroid Biochem Mol Biol. 154:120-129.
Merriman KE, Poindexter MB, Kweh MF, Santos JEP, Nelson CD. 2017. Intramammary dihydroxyvitamin D3 treatment increases expression of host-defense genes in mammary immune cells of lactating dairy cattle. J Steroid Biochem Mol Biol. 173:33-41.
Merriman KE, Powell JL, Santos JEP, Nelson CD. 2018. Intramammary 25-hydroxyvitamin D3 treatment modulates innate immune responses to endotoxin-induced mastitis. J Dairy Sci. 101:7593-7607.
Meyer MB, Pike JW. 2020. Mechanistic homeostasis of vitamin D metabolism in the kidney through reciprocal modulation of Cyp27b1 and Cyp24a1 expression. Steroid Biochem Mol Biol. 196:105500.
Naderi S, Bohlouli M, Yin T, König S. 2018. Genomic breeding values, SNP effects and gene identification for disease traits in cow training sets. Anim Genet. 49:178-192.
National Research Council. 2001. Nutrient requirements of dairy cattle. 7th rev. ed. Washington D.C: National Academy Press.
Nelson CD, Lippolis JD, Reinhardt TA, Sacco RE, Powell JL, Drewnoski ME, O’Neil M, Beitz DC, Weiss WP. 2016. Vitamin D status of dairy cattle: outcomes of current practices in the dairy industry. J Dairy Sci. 99:10150-10160.
Nelson CD, Nonnecke BJ, Reinhardt TA, Waters WR, Beitz DC, Lippolis JD. 2011. Regulation of mycobacterium-specific mononuclear cell responses by 25-hydroxyvitamin D3. PLoS One. 6: e21674.
Nelson CD, Reinhardt TA, Beitz DC, Lippolis JD. 2010a. In vivo activation of the intracrine vitamin D pathway in innate immune cells and mammary tissue during a bacterial infection. PLoS One. 5:e15469.
Nelson CD, Reinhardt TA, Lippolis JD, Sacco RE, Nonnecke BJ. 2012. Vitamin D signaling in the bovine immune system: a model for understanding human vitamin D requirements. Nutrients. 4:181-196.
Nelson CD, Reinhardt TA, Thacker TC, Beitz DC, Lippolis JD. 2010b. Modulation of the bovine innate immune response by production of 1alpha,25-dihydroxyvitamin D3 in bovine monocytes. J Dairy Sci. 93:1041-1049.
Nemere I, Garbi N, Hammerling G, Hintze KJ. 2012. Role of the 1,25D3-MARRS receptor in the 1,25(OH)2D3-stimulated uptake of calcium and phosphate in intestinal cells. Steroids. 77:897-902.
Nonnecke BJ, Waters WR, Foote MR, Horst RL, Fowler MA, Miller BL. 2003. In vitro effects of dihydroxyvitamin D3 on interferon-gamma and tumor necrosis factor-alpha secretion by blood leukocytes from young and adult cattle vaccinated with mycobacterium bovis BCG. Int J Vitam Nutr Res. 73:235-244.
Norman AW. 2012. The history of the discovery of vitamin D and its daughter steroid hormone. Ann Nutr Metab. 61:199-206.
Nykjaer A, Fyfe JC, Kozyraki R, Leheste JR, Jacobsen C, Nielsen MS, Verroust PJ, Aminoff M, de La Chapelle A, Moestrup SK, et al. 2001. Cubilin dysfunction causes abnormal metabolism of the steroid hormone 25(OH) vitamin D3. Proc Natl Acad Sci U S A. 98:13895-13900.
O’Brien MB, Beynon CL, McLoughlin RM, Meade KG. 2021. The immune response in bovine primary dermal fibroblasts is influenced by interleukin 8 promoter haplotype and vitamin D. Vet Immunol Immunopathol. 238:110291.
Oberg F, Botling J, Nilsson K. 1993. Functional antagonism between vitamin D3 and retinoic acid in the regulation of CD14 and CD23 expression during monocytic differentiation of U-937 cells. J Immunol. 150:3487-3495.
Peelen E, Knippenberg S, Muris AH, Thewissen M, Smolders J, Tervaert JW, Hupperts R, Damoiseaux J. 2011. Effects of vitamin D on the peripheral adaptive immune system: a review. Autoimmun Rev. 10:733-743.
Poindexter MB, Kweh MF, Zimpel R, Zuniga J, Lopera C, Zenobi MG, Jiang Y, Engstrom M, Celi P, Santos JEP, et al. 2020. Feeding supplemental 25-hydroxyvitamin D3 increases serum mineral concentrations and alters mammary immunity of lactating dairy cows. J Dairy Sci. 103:805-822.
Powe CE, Ricciardi C, Berg AH, Erdenesanaa D, Collerone G, Ankers E, Wenger J, Karumanchi SA, Thadhani R, Bhan I. 2011. Vitamin D-binding protein modifies the vitamin D-bone mineral density relationship. J Bone Miner Res. 26:1609-1616.
Prabhu AV, Luu W, Sharpe LJ, Brown AJ. 2017. Phosphorylation regulates activity of 7-dehydrocholesterol reductase (DHCR7), a terminal enzyme of cholesterol synthesis. J Steroid Biochem Mol Biol. 165:363-368.
Pulendran B, Tang H, Manicassamy S. 2010. Programming dendritic cells to induce T(H)2 and tolerogenic responses. Nat Immunol. 11:647-655.
Ramagopalan SV, Heger A, Berlanga AJ, Maugeri NJ, Lincoln MR, Burrell A, Handunnetthi L, Handel AE, Disanto G, Orton SM, et al. 2010. Knight JC.A ChIP-seq defined genome-wide map of vitamin D receptor binding: associations with disease and evolution. Genome Res. 20:1352-1360.
Reboul E, Goncalves A, Comera C, Bott R, Nowicki M, Landrier JF, Jourdheuil-Rahmani D, Dufour C, Collet X, Borel P. 2011. Vitamin D intestinal absorption is not a simple passive diffusion: evidences for involvement of cholesterol transporters. Mol Nutr Food Res. 55:691-702.
Reichel H, Koeffler HP, Barbers R, Norman AW. 1987. Regulation of 1,25-dihydroxyvitamin D3 production by cultured alveolar macrophages from normal human donors and from patients with pulmonary sarcoidosis. J Clin Endocrinol Metab. 65:1201-1209.
Reid IR, Baldock PA, Cornish J. 2018. Effects of leptin on the skeleton. Endocr Rev. 39:938-959.
Reid D, Toole BJ, Knox S, Talwar D, Harten J, O’Reilly DS, Blackwell S, Kinsella J, McMillan DC, Wallace AM. 2011. The relation between acute changes in the systemic inflammatory response and plasma 25-hydroxyvitamin D concentrations after elective knee arthroplasty. Am J Clin Nutr. 93:1006-1011.
Rodney RM, Martinez N, Block E, Hernandez LL, Celi P, Nelson CD, Santos EP, Lean IJ. 2018. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Vitamin D, mineral, and bone metabolism. J Dairy Sci 101:2519-2543.
Roizen JD, Casella A, Lai M, Long C, Tara Z, Caplan I, O’Lear L, Levine MA. 2018a. Decreased serum 25-hydroxyvitamin D in aging male mice is associated with reduced hepatic Cyp2r1 abundance. Endocrinology. 159:3083-3089.
Roizen JD, Li D, O’Lear L, Javaid MK, Shaw NJ, Ebeling PR, Nguyen HH, Rodda CP, Thummel KE, Thacher TD, et al. 2018b. CYP3A4 mutation causes vitamin D-dependent rickets type 3. J Clin Invest. 128:1913-1918.
Roizen JD, Long C, Casella A, O’Lear L, Caplan I, Lai M, Sasson I, Singh R, Makowski AJ, Simmons R, et al. 2019. Obesity decreases hepatic 25-hydroxylase activity causing low serum 25-hydroxyvitamin D. J Bone Miner Res. 34:1068-1073.
Rybchyn MS, Abboud M, Puglisi DA, Gordon-Thomson C, Brennan-Speranza TC, Mason RS, Fraser DR. 2020. Skeletal muscle and the maintenance of vitamin D status. Nutrients. 12:3270.
Safadi FF, Thornton P, Magiera H, Hollis BW, Gentile M, Haddad JG, Liebhaber SA, Cooke NE. 1999. Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J Clin Invest. 103:239-251.
Saito A, Iino N, Takeda T, Gejyo F. 2007. Role of megalin, a proximal tubular endocytic receptor, in calcium and phosphate homeostasis. Ther Apher Dial. 11:S23-26.
Saponaro F, Saba A, Zucchi R. 2020. An update on vitamin D metabolism. Int J Mol Sci. 21:6573.
Sassi F, Tamone C, D’Amelio P. 2018. Vitamin D: nutrient, hormone, and immunomodulator. Nutrients. 10:1656.
Schauber J, Dorschner RA, Coda AB, Büchau AS, Liu PT, Kiken D, Helfrich YR, Kang S, Elalieh HZ, Steinmeyer A, et al. 2007. Injury enhances TLR2 function and antimicrobial peptide expression through a vitamin D-dependent mechanism. J Clin Invest. 117:803-811.
Scherberich JE, Kellermeyer M, Ried C, Hartinger A. 2005. 1-alpha-calcidol modulates major human monocyte antigens and toll-like receptors TLR 2 and TLR4 in vitro. Eur J Med Res. 10:179-182.
Schlegel G, Ringseis R, Windisch W, Schwarz FJ, Eder K. 2012. Effects of a rumen-protected mixture of conjugated linoleic acids on hepatic expression of genes involved in lipid metabolism in dairy cows. J Dairy Sci. 95:3905-3918.
Schröder B, Breves G. 2006. Mechanisms and regulation of calcium absorption from the gastrointestinal tract in pigs and ruminants: comparative aspects with special emphasis on hypocalcemia in dairy cows. Anim Health Res Rev. 7:31-41.
Schwartz JB, Gallagher JC, Jorde R, Berg V, Walsh J, Eastell R, Evans AL, Bowles S, Naylor KE, Jones KS. 2018. Determination of free 25(OH)D concentrations and their relationships to total 25(OH)D in multiple clinical populations. J Clin Endocrinol Metab. 103:3278-3288.
Schwartz JB, Kane L, Bikle D. 2016. Response of vitamin D concentration to vitamin D3 administration in older adults without sun exposure: a randomized double-blind trial. J Am Geriatr Soc. 64:65-72.
Shieh A, Chun RF, Ma C, Witzel S, Meyer B, Rafison B, Swinkels L, Huijs T, Pepkowitz S, Holmquist B, et al. 2016. Effects of high-dose vitamin D2 versus D3 on total and free 25-hydroxyvitamin D and markers of calcium balance. J Clin Endocrinol Metab. 101:3070-3080.
Shieh A, Ma C, Chun RF, Wittwer-Schegg J, Swinkels L, Huijs T, Wang J, Donangelo I, Hewison M, Adams JS. 2018. Associations between change in total and free 25-hydroxyvitamin D with 24,25-dihydroxyvitamin D and parathyroid hormone. J Clin Endocrinol Metab. 103:3368-3375.
Silver J, Russell J, Sherwood LM. 1985. Regulation by vitamin D metabolites of messenger ribonucleic acid for preproparathyroid hormone in isolated bovine parathyroid cells. Proc Natl Acad Sci U S A. 82:4270-4273.
Sly LM, Lopez M, Nauseef WM, Reiner NE. 2001. 1alpha,25-Dihydroxyvitamin D3-induced monocyte antimycobacterial activity is regulated by phosphatidylinositol 3-kinase and mediated by the NADPH-dependent phagocyte oxidase. J Biol Chem. 276:35482-35493.
Small AG, Harvey S, Kaur J, Putty T, Quach A, Munawara U, Perveen K, McPhee A, Hii CS, Ferrante A. 2021. Vitamin D upregulates the macrophage complement receptor immunoglobulin in innate immunity to microbial pathogens. Commun Biol. 25:401.
Solanki SS, Singh P, Kashyap P, Sansi MS, Ali SA. 2021. Promising role of defensins peptides as therapeutics to combat against viral infection. Microb Pathog. 155:104930.
Sommerfeldt JL, Horst RL, Littledike TE, Beitz DC. 1979. In vitro degradation of cholecalciferol in rumen fluid. J Dairy Sci. 64:157. (Abstract).
Sommerfeldt JL, Napoli JL, Littledike ET, Beitz DC, Horst RL. 1983. Metabolism of orally administered [3H]ergocalciferol and [3H]cholecalciferol by dairy calves. J Nutr. 113:2595-2600.
Sordillo LM. 2016. Nutritional strategies to optimize dairy cattle immunity. J Dairy Sci. 99:4967-4982.
Tabaei S, Motallebnezhad M, Tabaee SS. 2021. Vitamin D Receptor ( VDR ) gene polymorphisms and risk of Coronary Artery Disease (CAD): systematic review and meta-analysis. Biochem Genet. 59:813-836.
Téllez-Pérez AD, Alva-Murillo N, Ochoa-Zarzosa A, Löpez-Meza JE. 2012. Cholecalciferol (vitamin D) differentially regulates antimicrobial peptide expression in bovine mammary epithelial cells: implications during Staphylococcus aureus internalization. Vet Microbiol. 160:91-98.
van de Peppel J, van Leeuwen JP. 2014. Vitamin D and gene networks in human osteoblasts. Front Physiol. 5:137.
Verboven C, Rabijns A, De Maeyer M, Van Baelen H, Bouillon R, De Ranter C. 2002. A structural basis for the unique binding features of the human vitamin D-binding protein. Nat Struct Biol. 9:131-136.
Vieira-Neto A, Poindexter MB, Nehme Marinho M, Zimpel R, Husnain A, Silva ACM, Prim JG, Nelson CD, Santos JEP. 2021. Effect of source and amount of vitamin D on function and mRNA expression in immune cells in dairy cows. J Dairy Sci. 104:10796-10811.
Vieth R, Kessler MJ, Pritzker KP. 1990. Species differences in the binding kinetics of 25-hydroxyvitamin D3 to vitamin D binding protein. Can J Physiol Pharmacol. 68:1368-1371.
Waldron JL, Ashby HL, Cornes MP, Bechervaise J, Razavi C, Thomas OL, Chugh S, Deshpande S, Ford C, Gama R. 2013. Vitamin D: a negative acute phase reactant. J Clin Pathol. 66:620-622.
Wang Y, Chen J, Lee CSD, Nizkorodov A, Riemenschneider K, Martin D, Hyzy S, Schwartz Z, Boyan BD. 2010. Disruption of Pdia3 gene results in bone abnormality and affects 1alpha,25-dihydroxy-vitamin D3-induced rapid activation of PKC. J Steroid Biochem Mol Biol. 121:257-260.
Wang X, Shapses SA, Al-Hraishawi H. 2017. Feee and bioavailable 25-hydroxyvitamin D levels in patients with primary hyperparathyroidism. Endocr Pract. 23:66-71.
Wasserman RH. 1975. Active vitamin D-like substances in Solanum malacoxylon and other calcinogenic plants. Nutr Rev. 33:1-5.
Waters WR, Nonnecke BJ, Rahner TE, Palmer MV, Whipple DL, Horst RL. 2001. Modulation of mycobacterium bovis-specific responses of bovine peripheral blood mononuclear cells by
dihydroxyvitamin D3. Clin Diagn Lab Immunol. 8:1204-1212.
Wen G, Eder K, Ringseis R. 2020. 1,25-hydroxyvitamin D3 decreases endoplasmic reticulum stress-induced inflammatory response in mammary epithelial cells. PLoS One. 15:e0228945.
Wilkens MR, Cohrs I, Lifschitz AL, Fraser DR, Olszewski K, Schröder B, Breves G. 2013. Is the metabolism of 25-hydroxyvitamin D3 age-dependent in dairy cows? J Steroid Biochem Mol Biol. 136:44-46.
Wilkens MR, Firmenich CS, Schnepel N, Muscher-Banse AS. 2019. A reduced protein diet modulates enzymes of vitamin D and cholesterol metabolism in young ruminants. J Steroid Biochem Mol Biol. 186:196-202.
Wilkens MR, Kunert-Keil C, Brinkmeier H, Schröder B. 2009. Expression of calcium channel TRPV6 in ovine epithelial tissue. Vet J. 182:294-300.
Wilkens MR, Muscher-Banse AS. 2020. Review: regulation of gastrointestinal and renal transport of calcium and phosphorus in ruminants. Animal. 14:s29-s43.
Williams MH, Van Alstyne EL, Galbraith RM. 1988. Evidence of a novel association of unsaturated fatty acids with Gc (vitamin D-binding protein). Biochem Biophys Res Commun. 153:1019-1024.
Wu J, Shang DP, Yang S, Fu DP, Ling HY, Hou SS, Lu JM. 2016. Association between the vitamin D receptor gene polymorphism and osteoporosis. Biomed Rep. 5:233-236.
Yu ZH, Chen M, Zhang QQ, Hu X. 2018. The association of vitamin d receptor gene polymorphism with lung cancer risk: an update meta-analysis. Comb Chem High Throughput Screen. 21:704-710.
Yue Y, Hym0ller L, Jensen SK, Lauridsen C. 2018. Effect of vitamin D treatments on plasma metabolism and immune parameters of healthy dairy cows. Arch Anim Nutr. 72:205-220.
Yue Y, Hym0ller L, Jensen SK, Lauridsen C, Purup S. 2017. Effects of vitamin D and its metabolites on cell viability and Staphylococcus aureus invasion into bovine mammary epithelial cells. Vet Microbiol. 203:245-251.
Zafalon RVA, Ruberti B, Rentas MF, Amaral AR, Vendramini THA, Chacar FC, Kogika MM, Brunetto MA. 2020. the role of vitamin D in small animal bone metabolism. Metabolites. 10:496.
Zasloff M. 2002. Antimicrobial peptides of multicellular organisms. Nature. 415:389-395.
Zella LA, Shevde NK, Hollis BW, Cooke NE, Pike JW. 2008. Vitamin D-binding protein influences total circulating levels of 1,25-dihydroxyvitamin D3 but does not directly modulate the bioactive levels of the hormone in vivo. Endocrinology. 149:3656-3667.
Zmijewski MA, Carlberg C. 2020. Vitamin D receptor(s): in the nucleus but also at membranes? Exp Dermatol. 29:876-884.
Some related items in VitaminDWiki
No tuberculosis if more than 80 ng of vitamin D (cattle) - Jan 2022
Dogs are 3.4X less likely to have Cancer if > 40ng of vitamin D
Companion animals (dog, cats) need vitamin D too – March 2016
Animals get a lot more vitamin D than minimum recommended for humans – 2016
Immunity category
Poor immune system associated with low Vitamin D (dogs in this case) – June 2020
Pigs used to get the flu only in the winter ( had got Vitamin D in the summer) - Nov 2018
Gene charts: this study and VitaminDWiki