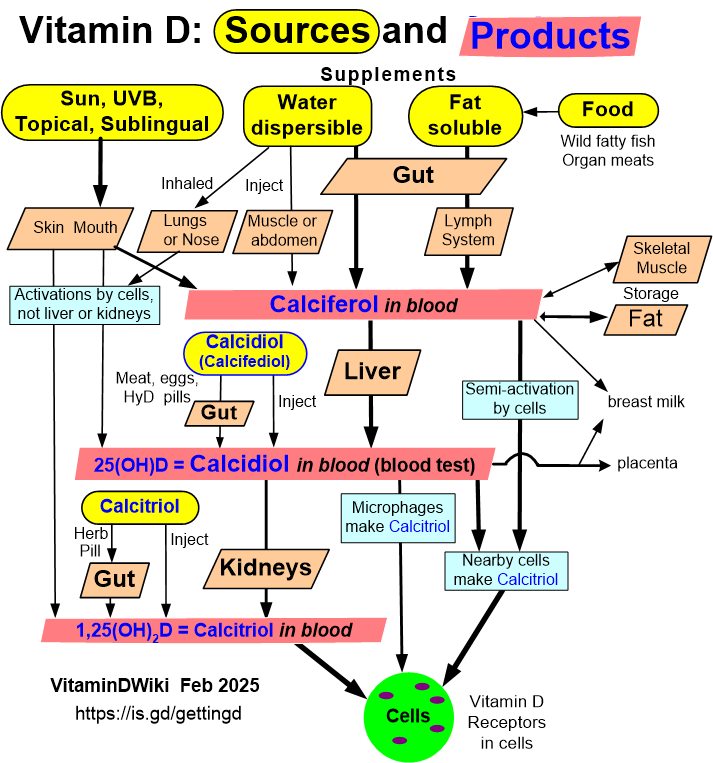
Calcifediol = semi-activated vitamin D = Calcidiol
Calcifediol: Mechanisms of Action
Nutrients 2023, 15, 4409. https:// doi.org/10.3390/nu15204409
Simone Donati1, Gaia Palmini 1 , Cinzia Aurilia 1, Irene Falsetti1, Francesca Marini 2 , Francesca Giusti 1 , Teresa Iantomasi1 and Maria Luisa Brandi 2,3,*@
Department of Experimental and Clinical Biomedical Sciences, University of Florence, 50139 Florence, Italy; simone.donati@unifi.it (S.D.); gaia.palmini@umfi.it (G.P.); cinzia.aurilia@umfi.it (C.A.); irene.falsetti@unifi.it (I.F.); francesca.giusti@unifi.it (F.G.); teresa.iantomasi@unifi.it (T.I.)
Fondazione Italiana Ricerca Sulle Malattie dell'Osso (FIRMO Onlus), 50129 Florence, Italy; francesca.marini@unifi.it
Donatello Bone Clinic, Villa Donatello Hospital, 50019 Sesto Fiorentino, Italy * Correspondence: marialuisa@marialuisabrandi.it
Due to its essential role in calcium and phosphate homeostasis, the secosteroid hormone calcitriol has received growing attention over the last few years. Calcitriol, like other steroid hormones, may function through both genomic and non-genomic mechanisms. In the traditional function, the interaction between the biologically active form of vitamin D and the vitamin D receptor (VDR) affects the transcription of thousands of genes by binding to repeated sequences present in their promoter region, named vitamin D-responsive elements (VDREs). Non-transcriptional effects, on the other hand, occur quickly and are unaffected by inhibitors of transcription and protein synthesis. Recently, calcifediol, the immediate precursor metabolite of calcitriol, has also been shown to bind to the VDR with weaker affinity than calcitriol, thus exerting gene-regulatory properties. Moreover, calcifediol may also trigger rapid non-genomic responses through its interaction with specific membrane vitamin D receptors. membrane - associated VDR (mVDR) and protein disulfide isomerase family A member 3 (Pdia3) are the best-studied candidates for mediating these rapid responses to vitamin D metabolites. This paper provides an overview of the calcifediol-related mechanisms of action, which may help to better understand the vitamin D endocrine system and to identify new therapeutic targets that could be important for treating diseases closely associated with vitamin D deficiency.
📄 Download the PDF from VitaminDWiki
Introduction
Vitamin D is found in virtually every form of life, from phytoplankton to humans, and is thought to be one of the oldest hormones on Earth [1]. Originally, vitamin D was defined as a vitamin because it could be obtained from food sources, mainly fatty fish, fish oils, dairy products, and some mushrooms.
Dietary vitamin D (vitamin D2 or D3) is normally absorbed by the small intestine along with other dietary fats [2]. The presence of fat in the intestinal lumen triggers the release of bile acids, which initiate emulsification and help to form lipid-containing micelles [3]. After ingestion, exogenous vitamin D is packaged into chylomicrons for transport to the liver. Part of the vitamin D contained in chylomicrons can be absorbed by adipose tissue and skeletal muscle [4]. Once the residual chylomicrons reach the liver, a vitamin D-binding protein (DBP) allows them to enter the liver cells, which then facilitates their transport to the various tissues that need them.
However, for humans, the photochemical production of 7-dehydrocholesterol (7-DHC) is the most important natural source of vitamin D that takes place in the basal layer of the epidermis in the skin [5]. In particular, the p-ring photodegradation of 7-DHC stimulated by solar ultraviolet type B (UVB) photon irradiation (around 280 to 320 nm) results in the
production of previtamin D3 that undergoes a thermal isomerization to form vitamin D3 (or cholecalciferol) or alternatively photoconverted ia two nonactive forms, such as tachys- terol and lumioterol [6,7]- Fohowing release from Uae cells, vitamin D3 enters circulation and is transported in titer circulation bound to a vitamin D-binding protein (DBP) to storage tissu es or the liver [s,9]. In the hepatocyte s, vitamin D3 is rapidly hydroxylated at the C-25 position by 25-hydroxylase, a cytochrome P450 enzyme (mainly the CYP2R1), thus producing calcitriol or calcidiol (25(OH)D3). Once synthesized, DBP-bounp 25(OH)D3 is secreted into the blood end transported to the? kidney to obtain the btologically active form calcitcto1 ( 1a,25(OH)2D3) (Figure 1).
The average plasma half-life of 25(OH)D3 ;s approximately between 20 and 214 days, the highest compered to other vitamin D metabolites [10]. Therefore, the measurement of 25(OH)D3 is the gold standard method to determine the body's vitamin D storage and status. To be exact, 25(OH)D3 is 1 a-hydroxylated by another CYP45a-dependent system activity (CYP27B1) in the mitochondria of the proximal convoluted tubule cells [ 11]. This reaction is closely regulated by blood phosphate and calcium levels through fibroblast: growth factor 23 (FGF-23) and parathyroid hormone (PTH). Calcitriol is involved in the regulation of plasma concentrations of ionized calcium and phosphato by modulating their renal excretion, intestinal absorption, and calcium bone mobilization. If levels of 1a,25(OH)2D3 rise, calcitriol induces its degradation by stimulating the expression of 24-hydroxylase (CYP24A1), which is also responsible for the catabolism of 25(OH)D3 [8,10]. Generally, the 24-hydroxylation reaction is followed by several oxidation reactions and sometimes the conjugation with glucuronic acid to form many compounds that are excreted through the bile [8]. Under low serum calcium conditions, the parathyroid glands secrete PTH, which stimulates the expression of 1a-hydroxylase, leading to enhanced 1a,25(OH)2D3 activation [12]. PTH also inhibits 24-hydroxylase and induces the synthesis of FGF-23 by osteoclasts and osteocytes, which acts to reduce the expression of renal sodium phosphate transporter [13,14]. FGF-23 can also regulate vitamin D homeostasis by inhibiting renal expression of 1a-hydroxylase and inducing 24-hydroxylase, thereby reducing serum calcitriol levels, which in turn reduce serum calcium levels in hyperphosphatemic conditions [15].
The human population is currently experiencing a high prevalence of moderate to severe vitamin D deficiency, which has detrimental effects on musculoskeletal and extra- skeletal systems. There is a general but not unanimous consensus that 25(OH)D3 levels <20 ng/mL are inadequate for maintaining musculoskeletal health, leading to rickets in children, osteomalacia in adults, and secondary hyperparathyroidism, while no serum 25(OH)D3 concentration is recommended for extra-skeletal health outcomes [16].
Based on more than 500 studies worldwide, 88% of the world's population have serum 25(OH)D3 levels <30 ng/mL, approximately 37% have values below 20 ng/mL, and about 7% have levels below 12 ng/mL [17]. Moreover, there are some groups or regions whose prevalence of vitamin D deficiency is higher.
The causes of vitamin D deficiency are varied and include limited sunlight exposure, sunscreen use, inadequate intake of foods containing vitamin D, dark skin pigmentation, patients with intestinal malabsorption syndromes, genetic diseases of vitamin D metabolizing enzymes or the vitamin D receptor (VDR), drugs that can interfere with the vitamin D absorption or metabolism, hepatic disease, obesity, aging, and renal disease [18,19]. On this latter aspect, approximately 80% of chronic kidney disease (CKD) patients have a high prevalence of vitamin D deficiency/insufficiency. Considering the extra-renal expression of 1a-hydroxylase, increasing interest was placed on vitamin D supplementation, including vitamin D2, vitamin D3, and 25(OH)D3, highlighting the importance of peripheral production of calcitriol. Despite the Italian Society of Nephrology (SIN) having already published a series of guidelines about vitamin D in CKD patients in 2016, indicating to supplement patients with CKD stages 3-5 and serum 25(OH)D3 concentration <30 ng/mL, the risk of toxicity should always be monitored, including adverse health effects associated with hypervitaminosis D, such as hyperphosphatasemia and hypercalcemia [20].
There are only a limited number of strategies to prevent or correct hypovitaminosis D. Increasing sun time exposure could, on the one hand, increase the skin synthesis of vitamin D, but on the other hand, considering that UVB is also photocarcinogen, it is not advisable to recommend a greater time exposure to sun, especially for those people with higher sensitivity to DNA [21]. In this light, it is well established that most dermatologists recommend avoiding long-term exposure to UVB light to prevent the development of long-term consequences, such as skin cancer. Furthermore, there are other limitations to implementing this strategy in real life, such as climatic circumstances, skin color, lifestyle, and cultural and religious habits [22]. Another option could be to increase the consumption of vitamin D-containing foods, but this is not feasible because there are no sufficient oils from fish in the ocean, one of the major dietary sources of vitamin D, to correct vitamin D deficiency status around the world. A good strategy is the intake of vitamin D- or calcifediol-fortified foods, and it has been successfully implemented in many northern European countries such as Finland [23]. However, this option becomes less effective in populations with a great variety of food preferences. Since the discovery of vitamin D, vitamin D supplementation has been a widely used valid strategy to prevent or correct such deficiency. The preferred compound globally used is vitamin D3, even though vitamin D2 is the vitamin D supplement of choice in the USA for the Steenbock patent and India because cholecalciferol is from animal origin [24,25]. Apart from these metabolites, even calcifediol has been considered as an oral vitamin D supplement to help people who are more likely to acquire hypovitaminosis D [26]. Based on Italian guidelines (https://www.aifa.gov.it/en/nota-96 (accessed on 29 September 2023)), the recommended dosage regimens for calcifediol and vitamin D3 depend on the individual's vitamin D status. In particular, an intake of 266 gg of calcifediol twice per month is recommended when serum 25(OH)D3 concentration is lower than 30 nmol/L. In a range of serum 25(OH)D3 concentration between 30 and 50 nmol/L, the recommended dosage for calcifediol is 266 gg once a month, while no vitamin D supplementation is generally not needed for those people with vitamin D blood levels above 50 nmol/L.
The main function of vitamin D is to regulate calcium-phosphate homeostasis, thereby contributing to maintaining proper bone health. The calciotropic hormone calcitriol functions as a steroid by binding to the intracellular VDR [27], a member of the steroid nuclear receptor superfamily, which includes retinoic acid, thyroid hormone, adrenal steroids, and sex hormones. This binding results in the modulation (suppression or activation) of gene expression [10,28-31].
Given that the presence of VDR is well documented in the cells of various organs, it has been established that the proper functioning of musculoskeletal, nervous, cardiovascular, and immune systems is strongly dependent on vitamin D [6,28]. Therefore, vitamin D deficiency status was not only described as a risk factor associated with the occurrence of rickets or osteoporosis [32] but also with the altered function of the immune system and the ability to mobilize a response to invading pathogens, including influenza and SARS-CoV-2 [33]. Over the last decade, it has been reported that vitamin D has a role in brain development and function [34]. Furthermore, it was suggested that levels of serum 25(OH)D3 in the range of 40 to 60 ng/mL are beneficial, reducing the occurrence and aggressiveness of several types of cancer [35] and showing cardioprotective [36] and neuroprotective [37] properties.
The biological effects of 1a,25(OH)2D3, like other steroid hormones, are mediated via both genomic and non-genomic mechanisms. Recently, 25(OH)D3 has been demonstrated to affect the expression of genes as well as to trigger non-genomic responses through its interaction with distinct membrane -associated, rapid-response steroid-binding receptors (MAARS) [22,38-40]. In this narrative review, we will provide an overview of the calcifediol- related mechanisms of action, which may help to better understand the vitamin D endocrine system and to identify new therapeutic targets that could be important for treating diseases closely associated with vitamin D deficiency.
Vitamin D Classical Actions: Regulation of Calcium and Phosphate Homeostasis
Calcitriol serves as one of the primary regulators of serum calcium levels within the optimal range by directly acting on three target tissues. Moreover, 1a,25(OH)2D3 suppresses parathyroid gene expression and parathyroid cell proliferation via the VDR, emphasizing its direct action on enhancing serum calcium levels [41].
One of the target organs is the intestine, where 1a,25(OH)2D3 stimulates intestinal calcium absorption [2]. This effect depends on the presence of dietary calcium, its intestinal solubility, and intestinal absorptive capacity as a result of the balance between transcellular and paracellular intestinal absorption. Transcellular transport involves three parts: calcium entry through specific calcium channels in the brush border membrane , intracellular transport through calbindin, and active calcium transport to the bloodstream on the basolateral surface, mainly via specific carriers.
The second organ is the kidney, in which both 1a,25(OH)2D3 and PTH promotes calcium reabsorption in the distal renal tubules [15]. Calcitriol affects (I) calcium entry through the apical membrane , (II) calcium diffusion mediated by calbamicin, and (III) active transport across the basolateral membrane . In addition, vitamin D inhibits phosphate reabsorption directly by inducing a-klotho and indirectly by enhancing FGF-23 osteocyte expression.
In the bones, 1a,25(OH)2D3 promotes the release of calcium from bone in a process requiring PTH [42]. In particular, the PTH-dependent calcitriol activation, as a consequence of the decreased serum calcium levels, stimulates the VDR-mediated formation and differentiation of osteoclasts. This activation results in the increase of skeletal calcium mobilization by stimulating the receptor activator for nuclear factor kappa-B ligand (RANKL) secretion, which, in turn, is a potent stimulator of osteoclastogenesis and bone resorption [43]. Moreover, vitamin D suppresses mineralization by enhancing pyrophosphate levels and osteopontin [44]. Therefore, vitamin D deficiency can lead to inadequate mineralization of the skeleton, thereby contributing to osteoporosis and fractures [45,46].
Vitamin D Molecular Actions
Genomic Response of Calcitriol
The secosteroid 1a,25(OH)2D3 is a calciotropic hormone that functions as a steroid molecule by its interaction with the intracellular VDR [27]. The VDR gene, which consists of eight coding exons, is found in fish, birds, and mammals and encodes a protein of 427 amino acid residues [47,48]. The two functional domains of the VDR are the conserved NH2-terminal DNA-binding domain named DBD and a highly variable COOH-terminal ligand-binding domain named LBD [22]. Calcitriol binding triggers conformational change, resulting in the dissociation of the repressor protein, thus promoting the dimerization of VDR to form either homodimers or heterodimers with retinoid X receptor (RXR) [10]. Once dimerized, both complexes (homo- and heterodimeric) bind to repeated sequences generally positioned in the physical proximity of transcription start site specifical of vitamin D-target genes named vitamin-D-responsive elements (VDREs) to regulate negatively or positively their expression [49]. Association of 1a,25(OH)2D3:VDR:RXR with the VDREs leads to the recruitment of coactivators that have histone acetylase activity, affecting the binding affinity of hystone proteins and DNA [50]. Furthermore, this large complex functions by recruiting RNA polymerase II enzyme to the transcription start site, thereby regulating the transcription of different genes involved in the control of calcium homeostasis (i.e., alkaline phosphatase (ALP), cytochrome P450 family (CYP450) 24 (CYP24), type I collagen (COL1A1), PTH, osteopontin, osteocalcin (bone gamma-carboxyglutamate protein (BGLAP), and transient receptor potential vanilloid type family member 6 (TRPV6)) [10,28-31,50]. The increase or decrease in protein expression levels caused by gene transcription regulation from steroid hormones is the result of their genomic actions. The timing of these actions is not immediate but rather delayed, as they take time for newly synthesized proteins and processing.
The totality of the above-mentioned transcriptional complexes defines the sensitivity and specificity of the various steroid hormones, including vitamin D [50]. In particular, the physiological response specificity is determined by the 1a,25(OH)2D3. The genetic specificity is guided by the VDRE. The cell or tissue specificity of the response depends on the different proteins recruited following the binding of 1a,25(OH)2D3-VDR-RXR to the VDRE. Finally, the biological response is caused by the activity of the vitamin D-responsive gene product.
25(OH)D3-Related Genomic Responses
Calcitriol is the only high-affinity (KD = 0.1 nM) ligand of the VDR [51], whereas the receptor affinity for 25(OH)D3 is 100- to 1000 times lower [52]. This discrepancy could be explained by the fact that 25(OH)D3 is missing a 1-OH-group, while generally the specific binding within the VDR's LBD pocket is achieved through interactions between a pair of polar amino acids and the three hydroxyl groups of 1,25(OH)2D3 (i.e., R274 and S237 bind the 1-OH-group, Y143 and S278 the 3-OH-group, and H305 and H397 the 25-OH-group) [53,54]. Nevertheless, the serum levels of 25(OH)D3 (50-250 nM) are 1000-fold higher compared to those of 1,25(OH)2D3 (0.05-0.15 nM), and therefore its levels should be enough for effective binding and to act as an agonistic VDR ligand [55,56].
As high levels of 25(OH)D3 are found in serum, blood cells are one the best experimental systems for investigating a possible gene regulatory function of 25(OH)D3. A study carried out by Hanel et al. [40] demonstrated that higher concentrations of 25(OH)D3, such as 1 pM and 10 pM nM, affect gene expression in peripheral blood mononuclear cells (PBMCs) isolated from five healthy individuals in a comparable way observed with 10 nM 1,25(OH)2D3. Among the 398 targets of 1 pM 25(OH)D3, approximately 86% also responded to 1,25(OH)2D3 in PBMCs, while the rate for the 477 target genes of 10 pM 25(OH)D3 was 78.0%. However, only two
genes, MYLIP and ABCG1, both involved in cholesterol transport, were specifically regulated by 10 gM 25(OH)D3.
In a subsequent study from the same research group [39], the authors found that the vitamin D metabolites 25(OH)D3 and 25(OH)D2 are equally able to directly modulate vitamin D-target gene expression, as well as 1,25(OH)2D3, the most potent VDR ligand, even though higher concentrations of 300 nM are required to observe these 25(OH)D-associated genomic responses.
Overall, these findings suggest that 25(OH)D3 within the physiological range (100-250 nM) does not affect gene expression, even though higher concentrations trigger such responses, although it is not to be excluded that the enzymatic formation of 1,25(OH)2D3 could partially contribute to modulate the transcriptome of PBMCs. This option cannot be excluded because these cells show reduced but significant expression levels of the CYP27B1 gene.
A similar expression pattern was also found in the prostate cancer cell line LNCaP [57].
The intriguing finding is that the metallothionein 2A gene was found to be a distinct target for 1,25(OH)2D3 but not for 25(OH)D3 [58]. Moreover, it is conceivable that the transcription factors encoded by genes activated by 1,25(OH)2D3 could, in turn, modulate the expression of additional gene sets [59], which could be defined as a secondary genomic response, even though this mechanism must still be defined even for 25(OH)D3.
Rapid Non-Genomic Actions
In 1942, Hans Selye described that progesterone showed an anesthetic effect immediately after peritoneum injection in rodents, which was different from what was observed with regard to its primary function, which occurred only within hours after its administration [60]. This is the first report of non-genomic actions mediated by steroid molecules. Then, Spach and Streeten showed that Na+ ions changed within a few minutes after the administration of aldosterone in dog erythrocytes, providing novel compelling evidence for the aldosterone-related non-genomic effects precisely for the absence of nuclei in these cells [61]. However, these non-transcriptional effects were not known until recently, when different rapid responses were recognized for various steroid hormones, including 1a,25(OH)2D3 [62].
In spite of the genomic counterpart, non-genomic rapid effects can be observed within seconds or minutes after stimulation, without the need for activation of gene expression and de novo subsequent protein synthesis. In this regard, these rapid mechanisms are not susceptible to molecules that inhibit the genomic effects, such as cycloheximide or actinomycin D, and also occur in response to steroids coupled to large proteins that do not allow their entry into the cells [29].
3.2.1. membrane -Associated Proteins and Targets for Vitamin D-Mediated Non-Genomic Responses
The idea of the existence of alternative non-genomic pathways activated by vitamin D was conceived by the pioneering studies of Nemere and colleagues in 1984 [63], who observed that 1a,25(OH)3D3 induced a rapid influx of intracellular calcium either by promoting its release from intracellular compartments or by improving the intestinal absorption in vascularly perfused duodenum from normal, vitamin D-replete chicks. This rapid influx of calcium within 14 min in response to 1a,25(OH)3D3 in a mechanism of genome activation- and protein synthesis-independent was named transcaltachia.
According to the time frame and the insensitivity of non-genomic responses to transcription and translation inhibitors, it was supposed that 1a,25(OH)2D3 triggers non-transcriptional effects by interacting with intracellular and membrane -associated macromolecules. This idea was corroborated by the observation that several secondary messengers were produced in response to 1a,25(OH)2D3, such as phosphatidylinositol (3,4,5)-trisphosphate (PIP3), calcium, and cyclic AMP (cAMP), culminating in a successful downstream activation of different protein kinases (PKs) (PKC, mitogen-activated protein (MAP) kinases, calcium/calmodulin-dependent protein kinase II gamma (CaMKIIG), and Src) [64-68]. Moreover, this secosteroid was also described to mediate the opening of calcium, chloride, and phosphate ion channels.
Several studies demonstrated that 1a,25(OH)2D3 interacts with the membrane isoform VDR (mVDR) associated with caveolin-1 (CAV1) and non-classical MAARS, resulting in fast non-genomic responses to vitamin D [69-71].
The existence of a distinct mVDR serving as the mediator of vitamin D-associated non- genomic signaling was described for the first time by the early studies of Norman et al. [69,70]. In this study, the authors observed that 1a,25(OH)2D3 in its 6-s-cis configuration was able to trigger rapid non-transcriptional responses, while the 6-s-trans configuration mediated the genomic responses.
Interestingly, the activation of at least some 1a,25(OH)2D3-induced non-genomic effects was displayed to be mVDR-dependent, such as the activation of SRC proto-oncogene non-receptor tyrosine kinase, and the regulation of several signaling pathways, such as Notch [72-74], sonic hedgehog (Shh) [75-80], and Wnt [81-84].
Another remarkable non-genomic activity of 1a,25(OH)2D3 was observed on models of adenosine diphosphate (ADP)- or collagen-induced platelet aggregation. Here, the authors [85] found that 1a,25(OH)2D3 had an inhibitory effect on platelet aggregation, which varied according to the state of diabetes. In particular, glycemic control was inversely and significantly associated with high platelet aggregation and reduced 25(OH)D3 levels. The absence of a nucleus and the presence of VDR in platelets indicated that this mechanism should be the outcome of the activation of non-genomic pathways.
Baran and colleagues [86] showed that 1a,25(OH)2D3 not only induced a rapid opening of calcium channels but also promoted a rapid activation of phospholipase C (PLC) in ROS 24/1 cells lacking VDR, indicating that these effects do not require the VDR involvement. The same finding was also validated in cultured costochondral chondrocytes derived from VDR-/- mice, where Boyan et al. [87] found that 1a,25(OH)2D3 was involved in the regulation of PKC activity.
This suggests that in addition to the presence of mVDR, the existence of other membrane receptors together with vitamin D could be essential for rapid non-genomic in response to 1a,25(OH)2D3.
One of the best characterized membrane -associated proteins was described in the above-mentioned study of Nemere et al. [63], who originally reported not only the ability of 1a,25(OH)2D3 to induce transcaltachia but also the purification of a plasmalemma receptor able to bind to the radiolabeled 1a,25(OH)2D3, with a KD-value of 0.72 nM. Later, this protein was termed protein disulfide isomerase family A member 3 (Pdia3) [88,89]. This protein interacts with two known molecular chaperones, calreticulin (CALR) and calnexin (CANX), playing a crucial role in the correct folding and export of newly synthesized glycoproteins [90]. Aside from cell membrane localization, Pdia 3 was also identified in mitochondria, cytosol, and nucleus, suggesting its involvement in different biological functions, such as cell protection from the ROS-induced damaging effects and the prevention of disorders associated with the accumulation of misfolded proteins [91]. The interaction between Pdia3 and 1a,25(OH)2D3 could also be an important protective mechanism against UV-driven DNA damage [92]. This response was associated with the activation of VDR-independent of the PKC signaling transduction pathway [93] and a rapid rise in intracellular calcium concentrations [94]. Further investigations have demonstrated that Pdia3-1a,25(OH)2D3 binding could be involved in the activation of PLA2 via PLA2- activating protein (PLAA) [71], MAPK1, and MAPK2 via the regulation of calcium/calmodulin-dependent protein kinase II gamma (CaMKIIG), PLA2, PLC, and PKC [65,95], and Wnt family member 5A (Wnt5A) [96].
Collectively, these findings pointed out that Pdia3 is essential for mediating the rapid non-genomics in response to vitamin D. Furthermore, some groups also suggested the nuclear localization of Pdia3, even though there is an ongoing debate about whether the secosteroid 1a,25(OH)2D3 could influence Pdia3 translocation into the nucleus and if the latter could act as a transcription factor or could promote the recruiting of other transcription factors [97].
25(OH)D3-Related Non-Genomic Responses
Calcifediol was, for a long time, recognized to be only a prohormone, a precursor of the biologically active form of vitamin D activated by the 1a-hydroxylation that takes place in proximal convoluted tubules of the kidney. As previously discussed in this review, recent evidence demonstrated that 25(OH)D3 interacts with VDR with a lower affinity (about 50 times) respect than 1a,25(OH)2D3.
Given the ability of 25(OH)D3 to join with the VDR, we [38] supposed that 25(OH)D3 could exert a fast non-genomic response, such as a sustained increase of intracellular calcium concentrations, in mesenchymal stem cells derived from human adipose tissue (hADMSCs), earlier depicted as an excellent cell model system for investigating the secosteroid hormone 1a,25(OH)2D3-related effects. We found out for the first time that 25(OH)D3 increases intracellular calcium levels in hADMSCs at much higher concentrations generally present in the human body (nanomolar range). As previously reported for the 25(OH)D3-related genomic effects, this could be the result of the reduced binding affinity of this vitamin D metabolite for VDR.
This finding was consistent with the non-genomic rapid effects of 25(OH)D3 in human spermatozoa, where Blomberg Jensen, M. et al. [98,99] showed that the immediate precursor of 1a,25(OH)2D3 was incapable of affecting calcium levels within the subnanomolar range, while a sustained but delayed intracellular calcium concentration rise was observed in human spermatozoa in response to of 25(OH)D3 at higher concentrations.
Interestingly, 25(OH)D3 was also found to regulate lipogenesis via a VDR-independent non-genomic effect that involved the processing and degradation of sterol regulatory element-binding protein (SREBP) cleavage-activating protein (SCAP) in the endoplasmic reticulum [100]. This intriguing observation could have a physiological significance in reducing the risk of metabolic syndrome-associated complications, in which an inverse correlation between serum 25(OH)D3 levels and severity was observed.
The disruption of endothelial integrity and the intensification of vascular ruptures have been shown to be implicated in pathogenic conditions, which can be prevented by adequate intake of vitamin D and its derivates. In addition to the canonical transcription-mediated vitamin D pathway, data showed that vitamin D3-related non-genomic responses could play an essential in maintaining both epithelial and endothelial cell stability [101]. Thus, vitamin D deficiency could impair the body's protective systems, resulting in the leakage of vascular fluid and worsening infections, ultimately leading to septicaemia [101]. Hence, the fast, non- transcriptional functions mediated by vitamin D might aid in the resolution of inflammation and infections and keep the endothelial junction integrity. As a result, deficiency in vitamin D could increase the vulnerability and the severity of infections and chronic diseases, which can lead to increased complications incidence rate and early death [102].
Discussion
The vitamin D endocrine system has been recognized as an essential part of the control of body calcium and phosphate levels, which facilitate adequate muscle performance, bone growth, and mineralization. These activities result from the activation of the VDR in intestinal, bone, and renal tissues by 1a,25(OH)3D3, primarily through direct interactions between 1a,25(OH)3D3-activated VDR and chromatin. Furthermore, it is now well-known that 1a,25(OH)2D3 has biological effects that do not involve gene transcription, which include the regulation of intracellular calcium as well as the activation of several signaling pathways via PKs and phosphatases [50].
More recently, studies showed that 25(OH)D3, the direct precursor of 1a,25(OH)2D3, can not only able to activate genomic responses but also modulate rapid non-genomic actions.
As described in this review, although the affinity of VDR for 25(OH)D3 is 100- to 1000-times lower compared to 1a,25(OH)2D3 [52], due both to a larger ligandbinding pocket and the absence of a 1-OH-group in the major circulating form of vitamin D, 25(OH)D3 is able to affect gene expression at concentrations higher than the physiological range. Moreover, a similar expression gene pattern was found between 25(OH)D3 and 1a,25(OH)2D3 either in the transcriptome of PBMCs and LNCaP [39,40,57]. Recently, it has been demonstrated that 1,25(OH)2D3 exerts regulatory effects on microRNA expression and long non-coding RNAs, potentially serving as a potential anticancer mechanism [103,104]. In this regard, no evidence has been established for the 25(OH)D3-induced regulation of these classes of non-coding RNA so far.
Furthermore, there is increasing evidence that 25(OH)D3 can also induce rapid, non- genomic activities, although their impact on physiological processes has not been clarified yet. In this regard, one of the possible physiological implications of the 25(OH)D3-mediated rapid non-genomic effect is the direct control of lipogenesis via a VDR-independent mechanism that involves the degradation of SCAP in the endoplasmic reticulum [100]. These findings could provide important clinical implications for alternative pathways activated by 25(OH)D3 to reduce the risk of complications associated with metabolic syndromes. Another clinical implication of 25(OH)D3-induced signaling pathways is the maintenance of epithelial and endothelial cell stability, which could be relevant for reducing the susceptibility and severity of chronic diseases associated with reduced levels of vitamin D. As reported in our previous study, 25(OH)D3 is able to increase intracellular calcium levels in hADMSCs here, similarly to what observed for the biologically active form of vitamin D. The rapid 1a,25(OH)2D3-induced stimulation of intestinal calcium is one of the most noticeable non-genomic effects of vitamin D, which serves to regulate the calcium level in the body [6,22]. However, additional studies are needed to provide conclusive evidence of the implications of such rapid action on meal feeding and calcium absorption physiology. Moreover, there is a growing body of evidence indicating that the rapid and non-genomic responses have the potential to both positively and negatively impact genomic functions [22]. This finding was demonstrated for 1a,25(OH)2D3, revealing that this secosteroid hormone can activate different signaling molecules involved in several signaling pathways, such as PI3K, PLC, and PLA2, thus affecting gene expression via primary regulatory elements of gene expression or using activated as a substrate. Furthermore, this interplay could influence both the effectiveness and the strength of gene regulation.
The field of vitamin D has made considerable advancements over the last years, expanding its understanding beyond the previously reported role in calcium regulation and rickets and osteomalacia prevention, respectively, in children and adults. Unfortunately, vitamin D-mediated non-genomic mechanisms are still not fully understood, and further studies are needed to reveal their functions in physiology, pathology, and clinical application potential. Although few studies are available in the literature on this field, a growing body of evidence has shown that 25(OH)D3 is an agonistic VDR ligand providing elucidation both for the direct gene regulatory properties and membrane non-genomic responses that have been defined for this vitamin D metabolite (Figure 2). Therefore, studies aiming to improve our understanding of the vitamin D endocrine system could be crucial to developing approaches for the prevention and treatment of chronic pathologies linked with suboptimal vitamin D status.
References
Carlberg, C. Vitamin D in the Context of Evolution. Nutrients 2022,14, 3018. [CrossRef] [PubMed]
Silva, M.C.; Furlanetto, T.W. Intestinal Absorption of Vitamin D: A Systematic Review. Nutr. Rev. 2018, 76, 60-76. [CrossRef] [PubMed]
Mulligan, G.B.; Licata, A. Taking Vitamin D with the Largest Meal Improves Absorption and Results in Higher Serum Levels of 25-Hydroxyvitamin D. J. Bone Min. Res. 2010, 25,928-930. [CrossRef] [PubMed]
Compston, J.E.; Merrett, A.L.; Hammett, F.G.; Magill, P. Comparison of the Appearance of Radiolabelled Vitamin D3 and 25-Hydroxy-Vitamin D3 in the Chylomicron Fraction of Plasma after Oral Administration in Man. Clin. Sci. 1981, 60, 241-243. [CrossRef]
Bikle, D.; Christakos, S. New Aspects of Vitamin D Metabolism and Action—Addressing the Skin as Source and Target. Nat. Rev. Endocrinol. 2020,16,234-252. [CrossRef]
Zmijewski, M.A. Nongenomic Activities of Vitamin D. Nutrients 2022,14, 5104. [CrossRef]
Cisneros, C.; Thompson, T.; Baluyot, N.; Smith, A.C.; Tapavicza, E. The Role of Tachysterol in Vitamin D Photosynthesis— A Non-Adiabatic Molecular Dynamics Study. Phys. Chem. Chem. Phys. 2017,19, 5763-5777. [CrossRef]
Jones, G. Pharmacokinetics of Vitamin D Toxicity. Am. J. Clin. Nutr. 2008, 88, 582S-586S. [CrossRef]
Bikle, D.D.; Schwartz, J. Vitamin D Binding Protein, Total and Free Vitamin D Levels in Different Physiological and Pathophysiological Conditions. Front. Endocrinol. 2019,10, 317. [CrossRef]
Gil, Ä.; Plaza-Diaz, J.; Mesa, M.D. Vitamin D: Classic and Novel Actions. Ann. Nutr. Metab. 2018, 72, 87-95. [CrossRef] [PubMed]
Norman, A.W. Sunlight, Season, Skin Pigmentation, Vitamin D, and 25-Hydroxyvitamin D: Integral Components of the Vitamin D Endocrine System. Am. J. Clin. Nutr. 1998, 67,1108-1110. [CrossRef]
Christakos, S. In Search of Regulatory Circuits That Control the Biological Activity of Vitamin D. J. Biol. Chem. 2017, 292,17559-17560. [CrossRef] [PubMed]
Nishimura, A.; Shinki, T.; Jin, C.H.; Ohyama, Y.; Noshiro, M.; Okuda, K.; Suda, T. Regulation of Messenger Ribonucleic Acid Expression of 1 Alpha,25-Dihydroxyvitamin D3-24-Hydroxylase in Rat Osteoblasts. Endocrinology 1994,134,1794-1799. [CrossRef] [PubMed]
Brown, R.B.; Razzaque, M.S. Dysregulation of Phosphate Metabolism and Conditions Associated with Phosphate Toxicity. Bonekey Rep. 2015, 4, 705. [CrossRef]
Perwad, F.; Portale, A.A. Vitamin D Metabolism in the Kidney: Regulation by Phosphorus and Fibroblast Growth Factor 23. Mol. Cell. Endocrinol. 2011, 347,17-24. [CrossRef]
Ramasamy, I. Vitamin D Metabolism and Guidelines for Vitamin D Supplementation. Clin. Biochem. Rev. 2020, 41, 103-126. [CrossRef]
Hilger, J.; Friedel, A.; Herr, R.; Rausch, T.; Roos, F.; Wahl, D.A.; Pierroz, D.D.; Weber, P.; Hoffmann, K. A Systematic Review of Vitamin D Status in Populations Worldwide. Br. J. Nutr. 2014,111, 23-45. [CrossRef]
Van Schoor, N.; Lips, P. Global Overview of Vitamin D Status. Endocrinol. Metab. Clin. N. Am. 2017, 46, 845-870. [CrossRef] [PubMed]
Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Endocrine Society Evaluation, Treatment, and Prevention of Vitamin D Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96,1911-1930. [CrossRef] [PubMed]
Cardoso, M.P.; Pereira, L.A.L. Native Vitamin D in Pre-Dialysis Chronic Kidney Disease. Nefrologia 2019, 39,18-28. [CrossRef] [PubMed]
Bouillon, R.; Quesada Gomez, J.M. Comparison of Calcifediol with Vitamin D for Prevention or Cure of Vitamin D Deficiency. J. Steroid Biochem. Mol. Biol. 2023, 228,106248. [CrossRef]
Donati, S.; Palmini, G.; Aurilia, C.; Falsetti, I.; Miglietta, F.; Iantomasi, T.; Brandi, M.L. Rapid Nontranscriptional Effects of Calcifediol and Calcitriol. Nutrients 2022,14,1291. [CrossRef] [PubMed]
Jääskeläinen, T.; Itkonen, S.T.; Lundqvist, A.; Erkkola, M.; Koskela, T.; Lakkala, K.; Dowling, K.G.; Hull, G.L.; Kröger, H.; Karppinen, J.; et al. The Positive Impact of General Vitamin D Food Fortification Policy on Vitamin D Status in a Representative Adult Finnish Population: Evidence from an 11-y Follow-up Based on Standardized 25-Hydroxyvitamin D Data. Am. J. Clin. Nutr. 2017,105,1512-1520. [CrossRef] [PubMed]
Jan, Y.; Malik, M.; Yaseen, M.; Ahmad, S.; Imran, M.; Rasool, S.; Haq, A. Vitamin D Fortification of Foods in India: Present and Past Scenario. J. Steroid Biochem. Mol. Biol. 2019,193,105417. [CrossRef] [PubMed]
Tripkovic, L.; Lambert, H.; Hart, K.; Smith, C.P.; Bucca, G.; Penson, S.; Chope, G.; Hyppönen, E.; Berry, J.; Vieth, R.; et al. Comparison of Vitamin D2 and Vitamin D3 Supplementation in Raising Serum 25-Hydroxyvitamin D Status: A Systematic Review and Meta-Analysis. Am. J. Clin. Nutr. 2012, 95,1357-1364. [CrossRef] [PubMed]
Donati, S.; Marini, F.; Giusti, F.; Palmini, G.; Aurilia, C.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. Calcifediol: Why, When, How Much? Pharmaceuticals 2023,16, 637. [CrossRef]
Di Rosa, M.; Malaguarnera, M.; Nicoletti, F.; Malaguarnera, L. Vitamin D3: A Helpful Immuno-Modulator. Immunology 2011, 134,123-139. [CrossRef]
Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and Human Health: Lessons from Vitamin D Receptor Null Mice. Endocr. Rev. 2008,29, 726-776. [CrossRef]
DeLuca, H.F. Evolution of Our Understanding of Vitamin D. Nutr. Rev. 2008, 66, S73-S87. [CrossRef]
Pike, J.W.; Meyer, M.B. The Vitamin D Receptor: New Paradigms for the Regulation of Gene Expression by 1,25-Dihydroxyvitamin D3. Endocrinol. Metab. Clin. N. Am. 2010, 39, 255-269. [CrossRef]
Gallieni, M.; Cozzolino, M.; Fallabrino, G.; Pasho, S.; Olivi, L.; Brancaccio, D. Vitamin D: Physiology and Pathophysiology. Int. J. Artif. Organs 2009, 32, 87-94. [CrossRef] [PubMed]
Charoenngam, N.; Ayoub, D.; Holick, M.F. Nutritional Rickets and Vitamin D Deficiency: Consequences and Strategies for Treatment and Prevention. Expert. Rev. Endocrinol. Metab. 2022,17, 351-364. [CrossRef] [PubMed]
Charoenngam, N.; Shirvani, A.; Holick, M.F. Vitamin D and Its Potential Benefit for the COVID-19 Pandemic. Endocr. Pract. 2021, 27, 484-493. [CrossRef] [PubMed]
Cui, X.; Gooch, H.; Petty, A.; McGrath, J.J.; Eyles, D. Vitamin D and the Brain: Genomic and Non-Genomic Actions. Mol. Cell. Endocrinol. 2017, 453,131-143. [CrossRef] [PubMed]
Carlberg, C.; Velleuer, E. Vitamin D and the Risk for Cancer: A Molecular Analysis. Biochem. Pharmacol. 2022, 196, 114735. [CrossRef] [PubMed]
Boucher, B.J.; Grant, W.B. Difficulties in Designing Randomised Controlled Trials of Vitamin D Supplementation for Reducing Acute Cardiovascular Events and in the Analysis of Their Outcomes. Int. J. Cardiol. Heart Vasc. 2020,29,100564. [CrossRef]
Moretti, R.; Morelli, M.E.; Caruso, P. Vitamin D in Neurological Diseases: A Rationale for a Pathogenic Impact. Int. J. Mol. Sci. 2018,19, 2245. [CrossRef] [PubMed]
Donati, S.; Palmini, G.; Romagnoli, C.; Aurilia, C.; Miglietta, F.; Falsetti, I.; Marini, F.; Zonefrati, R.; Galli, G.; Marcucci, G.; et al. In Vitro Non-Genomic Effects of Calcifediol on Human Preosteoblastic Cells. Nutrients 2021,13, 4227. [CrossRef]
Hanel, A.; Veldhuizen, C.; Carlberg, C. Gene-Regulatory Potential of 25-Hydroxyvitamin D3 and D2. Front. Nutr. 2022, 9, 910601. [CrossRef]
Hanel, A.; Bendik, I.; Carlberg, C. Transcriptome-Wide Profile of 25-Hydroxyvitamin D3 in Primary Immune Cells from Human Peripheral Blood. Nutrients 2021,13, 4100. [CrossRef]
Naveh-Many, T.; Marx, R.; Keshet, E.; Pike, J.W.; Silver, J. Regulation of 1,25-Dihydroxyvitamin D3 Receptor Gene Expression by 1,25-Dihydroxyvitamin D3 in the Parathyroid in Vivo. J. Clin. Investig. 1990, 86,1968-1975. [CrossRef]
Kuchuk, N.O.; van Schoor, N.M.; Pluijm, S.M.; Chines, A.; Lips, P. Vitamin D Status, Parathyroid Function, Bone Turnover, and BMD in Postmenopausal Women with Osteoporosis: Global Perspective. J. Bone Miner. Res. 2009, 24, 693-701. [CrossRef]
Harada, S.; Mizoguchi, T.; Kobayashi, Y.; Nakamichi, Y.; Takeda, S.; Sakai, S.; Takahashi, F.; Saito, H.; Yasuda, H.; Udagawa, N.; et al. Daily Administration of Eldecalcitol (ED-71), an Active Vitamin D Analog, Increases Bone Mineral Density by Suppressing RANKL Expression in Mouse Trabecular Bone. J. Bone Miner. Res. 2012, 27, 461-473. [CrossRef] [PubMed]
Murali, S.K.; Roschger, P.; Zeitz, U.; Klaushofer, K.; Andrukhova, O.; Erben, R.G. FGF23 Regulates Bone Mineralization in a 1,25(OH)2 D3 and Klotho-Independent Manner. J. Bone Miner. Res. 2016, 31,129-142. [CrossRef]
Nakamichi, Y.; Udagawa, N.; Suda, T.; Takahashi, N. Mechanisms Involved in Bone Resorption Regulated by Vitamin D. J. Steroid Biochem. Mol. Biol. 2018,177, 70-76. [CrossRef] [PubMed]
Courbebaisse, M.; Lanske, B. Biology of Fibroblast Growth Factor 23: From Physiology to Pathology. Cold Spring Harb. Perspect. Med. 2018, 8, a031260. [CrossRef]
DeLuca, H.F. Overview of General Physiologic Features and Functions of Vitamin D. Am. J. Clin. Nutr. 2004, 80,1689S-1696S. [CrossRef]
Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365-408. [CrossRef] [PubMed]
Feldman, D.; Malloy, P.J.; Gross, C. Chapter 9—Vitamin D: Biology, Action, and Clinical Implications. In Osteoporosis, 2nd ed.; Marcus, R., Feldman, D., Kelsey, J., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 257-303. ISBN 978-0-12-470862-4.
Morris, H.A. Vitamin D Metabolism and Molecular Modes of Action: New Insights into Vitamin D Activities. Med. Res. J. 2014, 2,1-5.
Haussler, M.R.; Haussler, C.A.; Bartik, L.; Whitfield, G.K.; Hsieh, J.-C.; Slater, S.; Jurutka, P.W. Vitamin D Receptor: Molecular Signaling and Actions of Nutritional Ligands in Disease Prevention. Nutr. Rev. 2008, 66, S98-S112. [CrossRef]
Lou, Y.-R.; Molnar, F.; Peräkylä, M.; Qiao, S.; Kalueff, A.V.; St-Arnaud, R.; Carlberg, C.; Tuohimaa, P. 25-Hydroxyvitamin D(3) Is an Agonistic Vitamin D Receptor Ligand. J. Steroid Biochem. Mol. Biol. 2010,118,162-170. [CrossRef] [PubMed]
Väisänen, S.; Ryhänen, S.; Saarela, J.T.A.; Peräkylä, M.; Andersin, T.; Mäenpää, P.H. Structurally and Functionally Important Amino Acids of the Agonistic Conformation of the Human Vitamin D Receptor. Mol. Pharmacol. 2002, 62, 788-794. [CrossRef]
Rochel, N.; Wurtz, J.M.; Mitschler, A.; Klaholz, B.; Moras, D. The Crystal Structure of the Nuclear Receptor for Vitamin D Bound to Its Natural Ligand. Mol. Cell 2000, 5,173-179. [CrossRef]
Wilhelm, F.; Mayer, E.; Norman, A.W. Biological Activity Assessment of the 26,23-Lactones of 1,25-Dihydroxyvitamin D3 and 25-Hydroxyvitamin D3 and Their Binding Properties to Chick Intestinal Receptor and Plasma Vitamin D Binding Protein. Arch. Biochem. Biophys. 1984, 233, 322-329. [CrossRef] [PubMed]
Gill, H.S.; Londowski, J.M.; Corradino, R.A.; Zinsmeister, A.R.; Kumar, R. Synthesis and Biological Activity of Novel Vitamin D Analogues: 24,24-Difluoro-25-Hydroxy-26,27-Dimethylvitamin D3 and 24,24-Difluoro-1 Alpha,25-Dihydroxy-26,27-Dimethylvitamin D3. J. Med. Chem. 1990, 33, 480-490. [CrossRef]
Ellfolk, M.; Norlin, M.; Gyllensten, K.; Wikvall, K. Regulation of Human Vitamin D(3) 25-Hydroxylases in Dermal Fibroblasts and Prostate Cancer LNCaP Cells. Mol. Pharmacol. 2009, 75,1392-1399. [CrossRef]
Susa, T.; Iizuka, M.; Okinaga, H.; Tamamori-Adachi, M.; Okazaki, T. Without 1 a-Hydroxylation, the Gene Expression Profile of 25(OH)D3 Treatment Overlaps Deeply with That of 1,25(OH)2D3 in Prostate Cancer Cells. Sci. Rep. 2018, 8, 9024. [CrossRef] [PubMed]
Warwick, T.; Schulz, M.H.; Günther, S.; Gilsbach, R.; Neme, A.; Carlberg, C.; Brandes, R.P.; Seuter, S. A Hierarchical Regulatory Network Analysis of the Vitamin D Induced Transcriptome Reveals Novel Regulators and Complete VDR Dependency in Monocytes. Sci. Rep. 2021,11,6518. [CrossRef] [PubMed]
Selye, H. Correlations between the Chemical Structure and the Pharmacological Actions of the Steroids. Endocrinology 1942, 30, 437-453. [CrossRef]
Spach, C.; Streeten, D.H.P. Retardation of Sodium Exchange in Dog Erythrocytes by Physiological Concentrations of Aldosterone, In Vitro. J. Clin. Investig. 1964, 43, 217-227. [CrossRef]
Schmidt, B.M.; Gerdes, D.; Feuring, M.; Falkenstein, E.; Christ, M.; Wehling, M. Rapid, Nongenomic Steroid Actions: A New Age? Front. Neuroendocr. 2000,21, 57-94. [CrossRef]
Nemere, I.; Yoshimoto, Y.; Norman, A.W. Calcium Transport in Perfused Duodena from Normal Chicks: Enhancement within Fourteen Minutes of Exposure to 1,25-Dihydroxyvitamin D3. Endocrinology 1984,115,1476-1483. [CrossRef]
Fleet, J.C. Rapid, membrane -Initiated Actions of 1,25 Dihydroxyvitamin D: What Are They and What Do They Mean? J. Nutr. 2004,134, 3215-3218. [CrossRef] [PubMed]
Doroudi, M.; Schwartz, Z.; Boyan, B.D. membrane -Mediated Actions of 1,25-Dihydroxy Vitamin D3: A Review of the Roles of Phospholipase A2 Activating Protein and Ca2+/Calmodulin-Dependent Protein Kinase II. J. Steroid Biochem. Mol. Biol. 2015, 147, 81-84. [CrossRef]
Dwivedi, P.P.; Hii, C.S.T.; Ferrante, A.; Tan, J.; Der, C.J.; Omdahl, J.L.; Morris, H.A.; May, B.K. Role of MAP Kinases in the 1,25-Dihydroxyvitamin D3-Induced Transactivation of the Rat Cytochrome P450C24 (CYP24) Promoter. Specific Functions for ERK1/ERK2 and ERK5. J. Biol. Chem. 2002,277, 29643-29653. [CrossRef] [PubMed]
Nutchey, B.K.; Kaplan, J.S.; Dwivedi, P.P.; Omdahl, J.L.; Ferrante, A.; May, B.K.; Hii, C.S.T. Molecular Action of 1,25- Dihydroxyvitamin D3 and Phorbol Ester on the Activation of the Rat Cytochrome P450C24 (CYP24) Promoter: Role of MAP Kinase Activities and Identification of an Important Transcription Factor Binding Site. Biochem. J. 2005, 389, 753-762. [CrossRef]
Dwivedi, P.; Gao, X.; Tan, J.; Evdokiou, A.; Ferrante, A.; Morris, H.; May, B.; Hii, C. A Role for the Phosphatidylinositol 3-Kinase-Protein Kinase C Zeta--Sp1 Pathway in the 1,25-Dihydroxyvitamin D3 Induction of the 25-Hydroxyvitamin D3 24-Hydroxylase Gene in Human Kidney Cells. Cell. Signal. 2010, 22, 543-552. [CrossRef] [PubMed]
Norman, A.W. Vitamin D Receptor: New Assignments for an Already Busy Receptor. Endocrinology 2006, 147, 5542-5548. [CrossRef] [PubMed]
Dormanen, M.C.; Bishop, J.E.; Hammond, M.W.; Okamura, W.H.; Nemere, I.; Norman, A.W. Nonnuclear Effects of the Steroid Hormone 1 Alpha,25(OH)2-Vitamin D3: Analogs Are Able to Functionally Differentiate between Nuclear and membrane Receptors. Biochem. Biophys. Res. Commun. 1994,201, 394-401. [CrossRef] [PubMed]
Zmijewski, M.A.; Carlberg, C. Vitamin D Receptor(s): In the Nucleus but Also at membrane s? Exp. Dermatol. 2020, 29, 876-884. [CrossRef]
Wang, H.; Wang, X.; Xu, L.; Zhang, J.; Cao, H. A Molecular Sub-Cluster of Colon Cancer Cells with Low VDR Expression Is Sensitive to Chemotherapy, BRAF Inhibitors and PI3K-mTOR Inhibitors Treatment. Aging 2019,11, 8587-8603. [CrossRef]
Olsson, K.; Saini, A.; Stromberg, A.; Alam, S.; Lilja, M.; Rullman, E.; Gustafsson, T. Evidence for Vitamin D Receptor Expression and Direct Effects of 1a,25(OH)2D3 in Human Skeletal Muscle Precursor Cells. Endocrinology 2016, 157, 98-111. [CrossRef] [PubMed]
Fuchs, E.; Raghavan, S. Getting under the Skin of Epidermal Morphogenesis. Nat. Rev. Genet. 2002, 3, 199-209. [CrossRef] [PubMed]
Bikle, D.D.; Jiang, Y.; Nguyen, T.; Oda, Y.; Tu, C. Disruption of Vitamin D and Calcium Signaling in Keratinocytes Predisposes to Skin Cancer. Front. Physiol. 2016, 7, 296. [CrossRef]
Bandera Merchan, B.; Morcillo, S.; Martin-Nuñez, G.; Tinahones, F.J.; Macías-González, M. The Role of Vitamin D and VDR in Carcinogenesis: Through Epidemiology and Basic Sciences. J. Steroid Biochem. Mol. Biol. 2017,167, 203-218. [CrossRef] [PubMed]
Hadden, M.K. Hedgehog and Vitamin D Signaling Pathways in Development and Disease. Vitam. Horm. 2016,100, 231-253. [CrossRef] [PubMed]
Lisse, T.S.; Saini, V.; Zhao, H.; Luderer, H.F.; Gori, F; Demay, M.B. The Vitamin D Receptor Is Required for Activation of cWnt and Hedgehog Signaling in Keratinocytes. Mol. Endocrinol. 2014,28,1698-1706. [CrossRef] [PubMed]
Teichert, A.E.; Elalieh, H.; Elias, P.M.; Welsh, J.; Bikle, D.D. Overexpression of Hedgehog Signaling Is Associated with Epidermal Tumor Formation in Vitamin D Receptor-Null Mice. J. Investig. Dermatol. 2011,131, 2289-2297. [CrossRef]
Teichert, A.; Elalieh, H.; Bikle, D. Disruption of the Hedgehog Signaling Pathway Contributes to the Hair Follicle Cycling Deficiency in Vdr Knockout Mice. J. Cell. Physiol. 2010, 225, 482-489. [CrossRef]
Tapia, C.; Suares, A.; De Genaro, P.; González-Pardo, V. In Vitro Studies Revealed a Downregulation of Wnt/|3-Catenin Cascade by Active Vitamin D and TX 527 Analog in a Kaposi's Sarcoma Cellular Model. Toxicol. In Vitr. 2020, 63,104748. [CrossRef]
Muralidhar, S.; Filia, A.; Nsengimana, J.; Pozniak, J.; O'Shea, S.J.; Diaz, J.M.; Harland, M.; Randerson-Moor, J.A.; Reichrath, J.; Laye, J.P.; et al. Vitamin D-VDR Signaling Inhibits Wnt/ |3-Catenin-Mediated Melanoma Progression and Promotes Antitumor Immunity. Cancer Res. 2019, 79, 5986-5998. [CrossRef]
Tang, L.; Fang, W.; Lin, J.; Li, J.; Wu, W.; Xu, J. Vitamin D Protects Human Melanocytes against Oxidative Damage by Activation of Wnt/P-Catenin Signaling. Lab. Investig. 2018, 98,1527-1537. [CrossRef]
Larriba, M.J.; González-Sancho, J.M.; Bonilla, F.; Muñoz, A. Interaction of Vitamin D with membrane -Based Signaling Pathways. Front. Physiol. 2014, 5, 60. [CrossRef]
Sultan, M.; Twito, O.; Tohami, T.; Ramati, E.; Neumark, E.; Rashid, G. Vitamin D Diminishes the High Platelet Aggregation of Type 2 Diabetes Mellitus Patients. Platelets 2019, 30,120-125. [CrossRef]
Baran, D.T.; Ray, R.; Sorensen, A.M.; Honeyman, T.; Holick, M.F. Binding Characteristics of a membrane Receptor That Recognizes 1 Alpha,25-Dihydroxyvitamin D3 and Its Epimer, 1 Beta,25-Dihydroxyvitamin D3. J. Cell. Biochem. 1994, 56, 510-517. [CrossRef] [PubMed]
Boyan, B.D.; Sylvia, V.L.; McKinney, N.; Schwartz, Z. membrane Actions of Vitamin D Metabolites 1alpha,25(OH)2D3 and 24R,25(OH)2D3 Are Retained in Growth Plate Cartilage Cells from Vitamin D Receptor Knockout Mice. J. Cell. Biochem. 2003, 90,1207-1223. [CrossRef] [PubMed]
Nemere, I.; Farach-Carson, M.C.; Rohe, B.; Sterling, T.M.; Norman, A.W.; Boyan, B.D.; Safford, S.E. Ribozyme Knockdown Functionally Links a 1,25(OH)2D3 membrane Binding Protein (1,25D3-MARRS) and Phosphate Uptake in Intestinal Cells. Proc. Natl. Acad. Sci. USA 2004,101, 7392-7397. [CrossRef] [PubMed]
Nemere, I.; Safford, S.E.; Rohe, B.; DeSouza, M.M.; Farach-Carson, M.C. Identification and Characterization of 1,25D3- membrane - Associated Rapid Response, Steroid (1,25D3-MARRS) Binding Protein. J. Steroid Biochem. Mol. Biol. 2004, 89-90, 281-285. [CrossRef] [PubMed]
Hettinghouse, A.; Liu, R.; Liu, C.-J. Multifunctional Molecule ERp57: From Cancer to Neurodegenerative Diseases. Pharmacol. Ther. 2018,181, 34-48. [CrossRef]
Mahmood, F.; Xu, R.; Awan, M.U.N.; Song, Y.; Han, Q.; Xia, X.; Zhang, J. PDIA3: Structure, Functions and Its Potential Role in Viral Infections. Biomed. Pharmacother. 2021,143,112110. [CrossRef] [PubMed]
Sequeira, V.B.; Rybchyn, M.S.; Tongkao-On, W.; Gordon-Thomson, C.; Malloy, P.J.; Nemere, I.; Norman, A.W.; Reeve, V.E.; Halliday, G.M.; Feldman, D.; et al. The Role of the Vitamin D Receptor and ERp57 in Photoprotection by 1 a,25-Dihydroxyvitamin D3. Mol. Endocrinol. 2012,26, 574-582. [CrossRef] [PubMed]
Khanal, R.; Nemere, I. membrane Receptors for Vitamin D Metabolites. Crit. Rev. Eukaryot. Gene Expr. 2007,17, 31-47. [CrossRef] [PubMed]
Yang, W.S.; Yu, H.; Kim, J.J.; Lee, M.J.; Park, S.-K. Vitamin D-Induced Ectodomain Shedding of TNF Receptor 1 as a Nongenomic Action: D3 vs D2 Derivatives. J. Steroid Biochem. Mol. Biol. 2016,155,18-25. [CrossRef] [PubMed]
Doroudi, M.; Chen, J.; Boyan, B.D.; Schwartz, Z. New Insights on membrane Mediated Effects of 1 a,25-Dihydroxy Vitamin D3 Signaling in the Musculoskeletal System. Steroids 2014, 81, 81-87. [CrossRef] [PubMed]
Doroudi, M.; Olivares-Navarrete, R.; Boyan, B.D.; Schwartz, Z. A Review of 1a,25(OH)2D3 Dependent Pdia3 Receptor Complex Components in Wnt5a Non-Canonical Pathway Signaling. J. Steroid Biochem. Mol. Biol. 2015,152, 84-88. [CrossRef] [PubMed]
Zhu, L.; Santos, N.C.; Kim, K.H. Disulfide Isomerase Glucose-Regulated Protein 58 Is Required for the Nuclear Localization and Degradation of Retinoic Acid Receptor Alpha. Reproduction 2010,139, 717-731. [CrossRef] [PubMed]
Blomberg Jensen, M.; Dissing, S. Non-Genomic Effects of Vitamin D in Human Spermatozoa. Steroids 2012, 77,903-909. [CrossRef] [PubMed]
Blomberg Jensen, M.; Bjerrum, P.J.; Jessen, T.E.; Nielsen, J.E.; Joensen, U.N.; Olesen, I.A.; Petersen, J.H.; Juul, A.; Dissing, S.; Jorgensen, N. Vitamin D Is Positively Associated with Sperm Motility and Increases Intracellular Calcium in Human Spermatozoa. Hum. Reprod. 2011,26,1307-1317. [CrossRef]
Asano, L.; Watanabe, M.; Ryoden, Y.; Usuda, K.; Yamaguchi, T.; Khambu, B.; Takashima, M.; Sato, S.-I.; Sakai, J.; Nagasawa, K.; et al. Vitamin D Metabolite, 25-Hydroxyvitamin D, Regulates Lipid Metabolism by Inducing Degradation of SREBP/SCAP. Cell Chem. Biol. 2017, 24,207-217. [CrossRef] [PubMed]
Bikle, D.D. Vitamin D Regulation of Immune Function. Vitam. Horm. 2011, 86,1-21. [CrossRef]
Ekwaru, J.P.; Zwicker, J.D.; Holick, M.F.; Giovannucci, E.; Veugelers, PJ. The Importance of Body Weight for the Dose Response Relationship of Oral Vitamin D Supplementation and Serum 25-Hydroxyvitamin D in Healthy Volunteers. PLoS ONE 2014, 9, e111265. [CrossRef] [PubMed]
Gallardo Martin, E.; Cousillas Castineiras, A. Vitamin D Modulation and microRNAs in Gastric Cancer: Prognostic and Therapeutic Role. Transl. Cancer Res. 2021,10, 3111-3127. [CrossRef] [PubMed]
Shahrzad, M.K.; Gharehgozlou, R.; Fadaei, S.; Hajian, P.; Mirzaei, H.R. Vitamin D and Non-Coding RNAs: New Insights into the Regulation of Breast Cancer. Curr. Mol. Med. 2021, 21,194-210. [CrossRef] [PubMed]
Analysis of the Efficacy and Safety of Weekly Calcifediol 100 µg in Vitamin D Deficient Patients
Jose Luis Pérez Castrillón 1ORCID,Esteban Jódar-Gimeno 2ORCID,Koldobika Molina 3,Aintzane García-Bea 4,Cristina Martínez Ostalé 3,* and Inmaculada Gilaberte 3
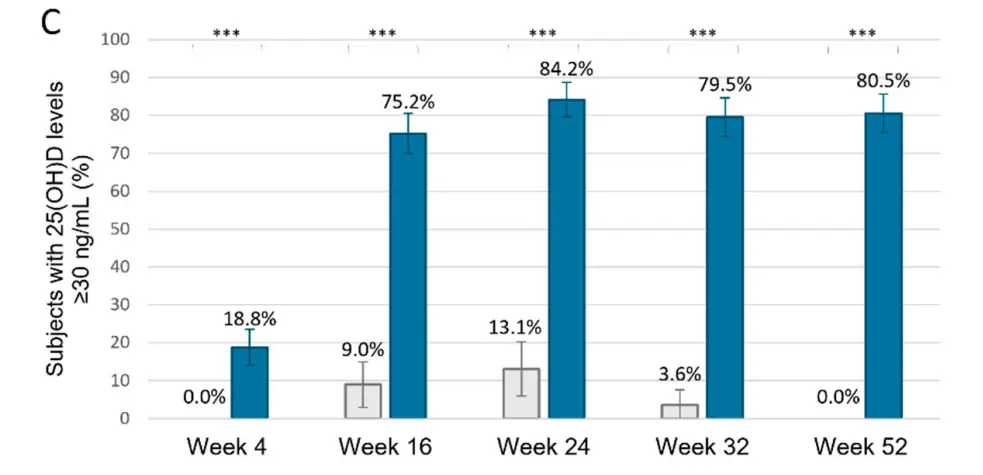
Pretty good response at 4 months
Background/Objectives: Adequate vitamin D levels are critical for overall health, yet vitamin D deficiency remains prevalent. This study aims to evaluate the efficacy and safety of a standardized weekly supplementation regimen of 100 μg calcifediol for patients with varying degrees of vitamin D deficiency.
Methods: A post hoc pool analysis was conducted from a randomized, double-blind, placebo-controlled, multicenter, two-cohort trial. Cohort 1 included vitamin D mild deficiency patients (25(OH)D levels > 10 < 20 ng/mL) and Cohort 2 severe deficiency patients (25(OH)D levels ≤ 10 ng/mL). As both had placebo and weekly calcifediol 100 μg arms (ratio 1:2), a pooled analysis of safety and efficacy was conducted. The primary outcome was the percentage of subjects achieving 25(OH)D levels ≥ 20 ng/mL and/or ≥30 ng/mL at various time points.
Results: A total of 401 participants across both cohorts were included in the analysis, 130 who received a placebo and 271 calcifediol 100 µg weekly. By week 52, 94.5% of individuals in the calcifediol group achieved 25(OH)D levels ≥ 20 ng/mL, compared to 25.3% in the placebo group (p < 0.0001). At this same week, 80.5% of subjects in the calcifediol group, but none in the placebo group (p < 0.0001), had 25(OH)D levels ≥ 30 ng/mL. The mean 25(OH)D level plateaued around 40.7 ng/mL from weeks 16 to 52. The frequency of treatment-emergent adverse events was similar in both groups, placebo and calcifediol.
Conclusions: Weekly supplementation of 100 μg calcifediol effectively restores vitamin D levels in individuals with both mild and severe deficiencies, demonstrating a favourable safety profile.
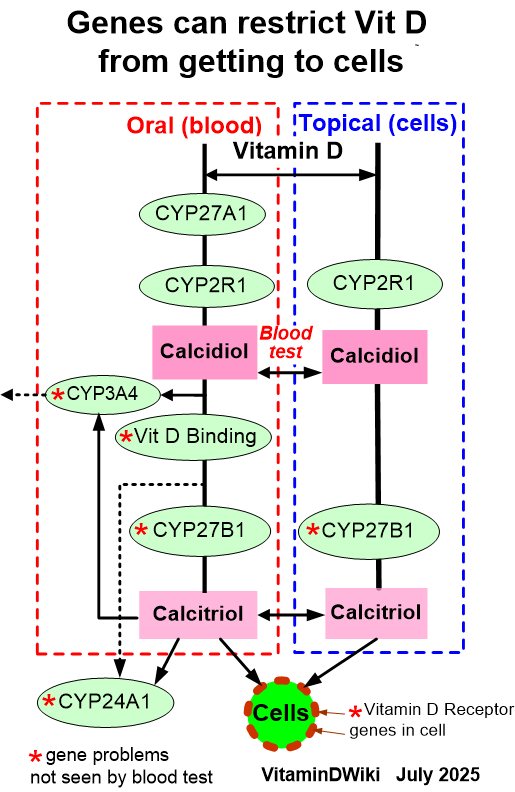
VitaminDWiki - Genetics chart shows the vitamin D genes
VitaminDWiki – Getting Vitamin D into your blood and cells has a chart: