Vitamin D and human function (from 24 years ago)
Diet and vitamin D: A review with an emphasis on human function
J. Nutr. Biochem. 5:58-65, 1994)
John J.B. Anderson and Svein U. Toverud
Department of Nutrition, Schools of Public Health and Medicine, and Department of Pharmacology, School of Medicine, University of North Carolina, Chapel Hill, NC USA
📄 Download the PDF from VitaminDWiki
Introduction
Vitamin D represents a unique molecule in human function because its requirements can be met from both the diet and skin photobiosynthesis. A mix of these two contributions is provided throughout the lifecycle, but the relative amounts of each may vary considerably, depending on the availability of vitamin D in the food supply and on the geographic latitude and atmospheric conditions. In recent decades, vitamin D fortification of dairy products and vitamin D supplementation have become important additional contributions to the total supply of this essential nutrient.
This review will cover the following topics: dietary sources, intakes and allowances of vitamin D through the lifecycle; age-related sun exposures and skin biosynthesis; circulating blood concentrations of vitamin D metabolites; important functional roles of the hormonal metabolite of vitamin D; age-related changes in vitamin D-driven calcium metabolism; modem biology of vitamin D; and human deficiency and toxicity.
Dietary sources, intakes, and allowances of vitamin D through the lifecycle
The major dietary sources of vitamin D in the USA, Canada, and a few other technologically advanced nations are fortified dairy products, especially milks and infant formulas, eggs, fortified margarines, and fishes, including their oils. In less advanced nations where the consumption of dairy products, eggs, and fish is minimal or non-existent, vitamin D intakes must be extremely low. For example, foods like cassava, maize (com), yam, plantain, rice, and other grains contain no vitamin D. Thus, subtropical and tropical peoples of the world must depend almost totally on skin biosynthesis to meet their tissue requirements of vitamin D. This section introduces terminology and usage of vitamin D molecules, and it provides data on intakes, allowances, and supplement usage of vitamin D.
Terminology of vitamin D molecules
Standard terminology of vitamin D molecules include the following: vitamin D, calciferol (general), animal—chole- calciferol (D3); fungal—ergocalciferol (D2): calcidiol, 25-hydroxycholecalciferolor25-OH-D3: calcitriol, 1,25-di- hydroxycholecalciferol or 1,25(OH)2D3, also vitamin D hormone; 24,25-dihydroxycholecalciferol or 24,25(OH)2D3 (low activity metabolite of unknown function).
Intakes of vitamin D
In the United States (USA), all processed cow’s milks are fortified with 10 p.g (400 International Units or IU) of cholecalciferol (or ergocalciferol) per quart (approximately 1 L), which exceeds by 10 fold the approximately 1 p,g per L of human breast milk. Actual estimated intakes by breast-feeding infants in the USA range between 0.5-1.5 |xg cholecalciferol a day and by formula-fed infants approximately 10 times higher for up to 6 months of age. Beyond 6 months, vitamin D intakes of infants and children relate directly to the amounts of fortified milks consumed. Two cups of milk (about 500 mL) provide approximately 5 p.g of calciferol (as either vitamin D2 or D3) per day. Other foods, of course, supply varying amounts of vitamin D. Adult consumption patterns of vitamin D-fortified dairy products are considerably lower than for infants and children, as assessed in a recent survey of the population of the USA. Intakes are approximately 2 |j.g cholecalciferol for males and 1.5 |xg for females.1 No other survey data on vitamin D intakes are available in the USA, but estimates of vitamin D intake by two different assessment methods in a study of African- American and white women in their mid-30s living in the New York City area yielded values of 140 to 150 IU per day.2
A wide variation in the vitamin D content of milks has recently been uncovered,3 because of an outbreak of vitamin D toxicity in the Boston, Massachusetts area.4 In addition, Holick et al.3 found that some skim milk samples contained virtually no vitamin D. These reports suggest that labeling of vitamin D-fortified dairy products throughout the USA could be highly inaccurate.
In the United Kingdom (UK), vitamin D intakes are lower than in the USA because milk fortification has been greatly reduced over the last few decades. The same is true for most of Europe. Scandinavian countries fortify margarine, but not dairy products.
In Japan, fish consumption provides almost 15% of total food energy and, thus, vitamin D intake from diverse fishes by adults is approximately 1 pg per day. Fortified milk products add an additional small amount of vitamin D to adult Japanese diets.
Estimates of vitamin D intakes by populations in the developing world are not available.
Allowances of vitamin D
Recommended intakes of vitamin D, which are based on both sunlight exposure for vitamin D skin biosynthesis and dietary sources, have been and remain extremely difficult to derive. In the US A, a set of recommended dietary allowances (RDA) has been established since the early 1940s. The most recent, tenth edition5 of the RDAs set forth RDAs according to specific ages and both genders (Table 1).
These most recent RDAs are the same as the allowances recommended in 1980 (ninth edition), at which time the RDA for young infants was lowered by about 25% from previous editions. The rationale for lowering the vitamin D RDA in 1980 was the fear of producing vitamin D toxicity from excessive intakes, as was found in the earlier UK experience with excessively fortified milk products.
Vitamin D allowances for other nations are generally lower than those of the USA, but higher intakes are recommended by nations, such as the former Soviet Union and Canada, where latitudes are significantly removed from the equator.
Supplements
In recent years, supplements of vitamin D have been increasingly recommended for the elderly, especially those who have become institutionalized or shut in because of limited mobility or confining conditions and diseases. Because the elderly have declining capabilities of skin biosynthesis and vitamin D conversion (see below), larger doses of vitamin D supplements are being recommended for this segment of the population. This recommendation is supported by low circulating concentrations of 25-OH-D6 or l,25(OH)2D.78 Such low blood levels of the 25-OH-vitamin D metabolite are now being defined as characteristic of vitamin D insufficiency rather than frank deficiency.
Table 1 Recommended dietary allowance of vitamin D, USA, 1989 5

Typical dosages of vitamin D supplements being recommended for elderly subjects in the USA are 400 IU per day or greater.
A recent report from Finland, where wintertime deficiencies of vitamin D among the elderly become relatively severe, demonstrated that intramuscular vitamin D injections of single yearly doses of 150,000 to 300,000 IU of ergocalcif- erol maintained serum levels of 25-OH-D within the normal range and reduced hip fracture incidence.9 Wintertime supplementation of elderly women in the northern latitude of the USA also resulted in improved 25-OH-D blood levels.10
Age-related sun exposures and skin biosynthesis
Energy from ultraviolet radiation (UV) derived from sunlight (and certain artificial lamps) can break the B-ring of 7- dehydrocholesterol in the epidermal layer of skin of humans and most other vertebrates to produce vitamin D , or cholecalciferol. The amount of skin biosynthesis per day is affected by many factors, such as latitude, sun (UV) exposure, dress, season of year, and skin melanin pigmentation (color). Webb et al.1 estimated that only 15 to 20 minutes exposure of arms and face at midday (c. 2 p.m.) will yield sufficient skin production of vitamin D sufficient to meet daily needs of adults living in Boston, Massachusetts, USA i.e., 5 ug (or 200 IU ). In the elderly, however, almost twice as much time is required to yield the same amount of vitamin D production by the skin.11 Stryd et al.12 and Lawson13 also demonstrated the significance of environmental conditions that can modify skin biosynthesis of vitamin D. Low skin production coupled with inadequate intake of vitamin D has been attributed as a primary cause of secondary hyperparathyroidism, osteomalacia, and hip fractures of elderly Chinese living in Hong Kong.14
Biotransformation of vitamin D and concentrations of circulating metabolites
The antirachitic factor, cholecalciferol (vitamin D3) is now considered a part of an endocrine system because it is synthesized in the skin under the influence of UV light and is ultimately converted in the kidney to a hormone, l,25(OH)2D (calcitriol), which circulates at low concentrations and exerts effects in many target organs.15
The precursor in the skin, 7-dehydrocholesterol (also called provitamin D), is converted to an intermediate form, previtamin D3, by opening the B-ring of the sterol molecule, a process mediated by UV light in the range 290-315 nm.1617 A subsequent rearrangement of the molecule takes place under the influence of skin temperature and leads to the formation of cholecalciferol, which is then released into the blood and binds to the circulating vitamin D-binding protein. In the elderly, the epidermal concentration of 7-dehydrocholesterol is diminished, and the rate of conversion to vitamin D3 is also reduced.18 When one considers that the elderly usually have reduced sun exposure as well, it becomes clear that they are at increased risk of developing vitamin D insufficiency unless they have an adequate dietary supply of vitamin D.18
The next biotransformation step occurs in the liver, where vitamin D3 or D2 is hydroxylated in the 25-position of the sterol by an enzyme reaction that is not closely controlled.19 In other words, the higher the blood concentration of vitamin D, the greater the amount of 25-OH-D produced. This metabolite circulates mostly bound to the vitamin D-binding protein. The concentration of this metabolite in blood is the best indicator of the adequacy of the supply of vitamin D, whether from the diet or the skin. The final biotransformation step occurs in the kidney, where 25-OH-D is hydroxylated further at the 1-position with formation of the active hormone, l,25(OH)2D. This hydroxylation is tightly regulated, in that the hydroxylase, a cytochrome P450 enzyme, is stimulated by elevated circulating parathyroid hormone concentrations, a fall in serum calcium (Ca), or a fall in the serum phosphate level. In the absence of any of these signals, 25-OH-D will be hydroxylated in the 24-position with the formation of 24,25 (OH)2D, a metabolite that lacks hormonal activity. Synthesis of l,25(OH)2D is inhibited by elevated circulating l,25(OH)2D levels and probably also by elevated serum Ca levels. In other words, the hormone is synthesized in response to changes reflecting either calcium or phosphate deficiency, or increased need.
In the presence of an adequate supply of the precursor, 25- OH-D, the circulating hormone level is increased during the pubertal period of rapid growth, and during the third trimester of pregnancy, as well as during lactation if milk production is high, as in mothers nursing twins. Alteration in the vitamin D endocrine system in old age, such as a decrease in the circulating l,25-(OH)2D level, has been implicated in the pathogenesis of postmenopausal osteoporosis because reduced serum l,25(OH)2D levels have been found in some women afflicted with this condition. However, there is no evidence indicating a general age-related decrease in serum l,25(OH)2D concentration. A recent study of a large group of healthy Caucasian adults ranging in age from 20-94 years revealed no decline in serum 25-OH-D or l,25(OH)2D concentrations in either sex.20 In another study, involving women between 26 and 88 years, serum l,25(OH)2D levels were actually higher after age 65 than in the youngest subjects, but the rate of intestinal Ca absorption was unaltered, suggesting decreased responsiveness to the hormone with increasing age followed by a compensatory increase in hormone synthesis.21
Circulating blood concentrations of major vitamin D metabolites
Blood concentrations of the major vitamin D metabolites, 25-OH-D and l,25(OH)2D have a fairly broad range in healthy individuals, but they do vary according to age, sex, and dietary intake or skin biosynthesis of vitamin D. Data reported in recent publications are emphasized in this review, especially those dealing with women and elderly subjects because of the important vitamin D-calcium relationship. While a seasonal variation exists for 25-OH-D, none has been found for l,25(OH)2D. Values of the major vitamin D metabolites are similar among African-American and white women in the USA.2
Table 2 provides data (means, SDs, and ranges) on circulating vitamin D metabolites by age, i.e., vitamin D status; these data are combined from reports of several different laboratories.
Vitamin D binding protein (DBP)
A hepatic-produced specific transport protein, DBP, binds large fractions of the circulating metabolites of vitamin D. The binding affinities of DBP for the metabolites are low, but the capacities are high. The qualitative order of affinities of DBP for vitamin D and its metabolites is as follows: 25- OH-D > 24,25(OH)2D > D > l,25(OH)2D.22 According to these authors, the binding affinity of DBP for vitamin D is approximately 300 times weaker than for 25-OH-D. As far as is known, DBP is never a limiting factor for carrying these metabolites in healthy individuals, even among the elderly with reasonably good liver function. DBP carries vitamin D or its metabolites from the fat depots, the major store of vitamin D, from skin after biosynthesis, and from the liver and kidney after biotransformations to other tissues. (Orally absorbed vitamin D is transported from the gut via chylomicrons.) The greatest quantity of a circulating metabolite carried by DBP is 25-OH-D, which, in effect, serves as the major readily available store of vitamin D. DBP, in effect, serves as a buffer against toxicity, because the free metabolites of vitamin, 25-OH-D and l,25(OH)2D, are toxigenic.
Oral therapy with vitamin D, calcitriol, and a calcitriol analog
Table 2__ Concentrations of major vitamin D metabolites across the lifecycle from reports in the literature
Therapeutic studies of elderly subjects using vitamin D have been conducted over the last two decades, but only one controlled investigation has been reported.23 Based on a review of these and several of his own reports Gallagher24 recommends the use of 800 IU of vitamin D per day for elderly individuals. In Japan, vitamin D [or l,25(OH)2D3 or its analog la-OH-D3] is favored as the therapy for osteoporotic patients.23 A recent study of osteoporotic women from New Zealand using low doses of calcitriol demonstrated a significant reduction in vertebral fractures after 3 years and no serious side effects .26
Oral therapy of 1 -a-hydroxy-cholecalciferol has recently been introduced in Japan and it has been shown to reduce the fracture rate of elderly subjects with low bone mass or fractures.27-28 The 1-a form must be converted in the liver to l,25(OH)2D by 25-hydroxylation. Dosages are generally low because of concerns about toxic effects. The efficacy of this therapy has not yet been fully evaluated.
Metabolic actions of l,25(OH)2D
It is generally agreed that all the actions attributed to vitamin D are mediated by the hormonal metabolite, l,25(OH)2D. An exception may be cases of vitamin D toxicity, when serum concentrations of 25-OH-D are increased many fold over the normal range; the 25-OH-D metabolite may then interact directly with the vitamin D receptor, mimic the actions of l,25(OH)2D, and cause hypercalcemia.19
Intestinal absorption
The most prominent action of l,25(OH)2D is the enhancement of intestinal absorption of both Ca and phosphate, an action that contributes to the maintenance of normal serum Ca and phosphate levels, which provides a supply of these ions for mineralization of bones and teeth. The component of intestinal Ca absorption stimulated by l,25(OH)2D is the active transport in the duodenum and to a lesser extent in the ileum. Such active transport, which takes place against a concentration gradient across the mucosal epithelium, is of greatest importance when the dietary Ca intake is low. There is ample evidence in experimental animals, but limited evidence in humans, that a reduced Ca intake leads to increased efficiency of Ca absorption, i.e., a relatively greater fraction of ingested Ca will be absorbed at low Ca intakes because of increased active transport mediated by an increase in serum 1,25(OH),D concentration.29 During periods of ample or high Ca intakes a relatively smaller fraction of the ingested Ca will be absorbed by active transport, and most of the Ca will be absorbed by passive transfer across the jejunum and the ileum. This process of adaptation of Ca absorption to the needs of the organism may not be equally efficient in all individuals and may require variable periods of time. In the elderly, this adaptational mechanism may be significantly reduced,30 but not all reports are in agreement with this finding.31
Bone tissue
The role of 1,25(OH)2D in bone metabolism is multifaceted. It is well documented that the presence of adequate amounts of the hormone is required for normal growth, maturation, and mineralization of bone, and of maintenance of mature bone tissue. As stated above, these effects of the hormone are largely due to the increased intestinal absorption of Ca and phosphate. However, osteoblasts and their precursors contain receptors for the hormone that mediate synthesis of a bone-specific protein, osteocalcin, that is associated with the bone formation process in a manner that is not yet clearly understood. In addition, some evidence exists that l,25(OH)2D stimulates the differentiation of osteoblasts.32 Also l,25(OH)2D indirectly stimulates the maturation and activity of osteoclasts. Osteoclasts do not have receptors for the hormone, but they respond to factors released from activated osteoblasts. High concentrations of the hormone have been shown to produce an increase in the number and activity of osteoclasts, which will result in an increased rate of bone resorption and net bone loss. The effect of the hormone on bone is therefore an increase in both rates of bone formation and bone resorption, i.e., an increase in bone turnover.33
Calcium homeostasis
A third action of l,25(OH)2D in bone is that of maintaining Ca homeostasis, i.e., the control of the Ca concentration in serum and extracellular fluid. The minute-to-minute and hour-to-hour regulation of the Ca concentration depends on the transfer of Ca from a pool of readily accessible Ca ions on the surface of the bone mineral crystals. This transfer appears to be under the control of a subpopulation of osteoblasts, and it is stimulated by l,25(OH)2D, as well as by parathyroid hormone. These hormones appear to function in concert even though they act by different mechanisms.
Other actions
Other effects of l,25(OH)2D include stimulation of renal conservation of Ca and phosphate, effects that are not well documented in humans.34 An interaction with the immune system involves activation of T-lymphocytes. However, the significance of this interaction is not clear, because vitamin D-deficient patients have little if any signs of impaired immunity.32 The hormonal form of vitamin D is also involved in the function of skeletal muscles, as evidenced by the presence of l,25(OH)2D receptors and reports of muscular weakness in patients with severe rickets.19 35 In the skin l,25(OH)2D promotes terminal differentiation of keratino- cytes. This action is the basis for the experimental use of topical application of the hormone in patients with the hyper- proliferative skin disease, psoriasis.36
Solar ultraviolet radiation exposure, and thereby vitamin D, has been suggested by epidemiologic analysis to protect against prostate cancer37 and possibly other cancers.32
Summary
In summary, l,25(OH)2D exerts important roles in intestinal absorption of Ca and phosphate, bone metabolism and mineralization, and Ca homeostasis. Other roles are not yet well understood and include conservation of Ca and phosphate by the kidney and interactions of the hormone with the immune system, skin, and skeletal muscle.
Mechanisms of action of l,25(OH)2D
Actions of the hormonal form of vitamin D fall under two categories, genomic and nongenomic.
Genomic action
The specific receptorfor 1,25(OH)2D that mediates the genomic action of the hormone, referred to as the vitamin D receptor (VDR), is a protein of approximately 48,000 molecular weight.38 It is present in specific cells in most tissues of the body,35 and is localized primarily in the nucleus, but possibly in the cytoplasm as well.39 The VDR has a specific domain responsible for binding the hormone and another domain that binds the hormone-receptor complex to DNA. The latter binding leads to transcription of specific proteins such as the osteocalcin in osteoblasts and the Ca-binding protein, calbindin D-9k, in the mucosal cells of the small intestine. Calbindin levels are closely associated with the rate of active Ca transport, and calbindin is believed to be involved in the transcellular movement of Ca.40 The calbindin D-9k gene of rats contains a l,25(OH)2D-responsive element that mediates the increased expression of this gene by 1,25(OH)2D in the intestine.41 The VDR studies revealed that l,25(OH)2D acts in the same manner as other steroid hormones. The clinical significance of the receptor studies is illustrated by the discovery of defects in the VDR in patients with hereditary vitamin D-dependent rickets type 2, a rare autosomal recessive condition. These defects, which impair binding of the receptor to the hormone and the binding of the complex to DNA, explain why the patients are unresponsive to both the endogenous hormone as well as to treatment with high doses of the hormone.
Nongenomic action
Several lines of evidence indicate that the vitamin D hormone can mediate changes in cells within seconds or a few minutes by an unknown mechanism. These changes, which include alteration in intracellular pH and Ca concentration, occur too rapidly to be explained by gene activation and new protein synthesis, and the actions of the hormone underlying these effects are therefore called nongenomic. Of particular interest is the observation that increased absorption of Ca from chick intestine can be observed as early as 5 minutes after administration of the hormone, a phenomenon called transcaltachia (rapid transfer of Ca).42 In rat colonic epithelial cells, l,25(OH)2D3 caused formation of diacylglycerol and 1,4,5-triphosphate, subsequent activation of protein kinase C, and an increase in intracellular calcium concentration; all these events occurred within seconds to minutes.43 Rapid effects of this vitamin D metabolite on phosphoinosi- tide metabolism were also detected in rat duodenal mucosal cells, but not prior to the time of development of the nuclear receptor by young suckling rat pups, which suggests the involvement of cell surface receptors that are similar or identical to the nuclear receptor.44 Data published by Norman et al.,45 however, indicate that the postulated D-hormone receptor in the cell membrane is different from the nuclear receptor. Based on studies of the hormone-induced transcription of the osteocalcin gene in osteoblast-like cells in culture, Baran et al.46 suggest that the rapid effect of l,25(OH)2D, serves to facilitate the genomic action. The physiological significance of the nongenomic action of the hormone in different cell types remains to be established.
Summary
While all the classical actions of calcitriol appear to be explained satisfactorily by a genomic mechanism, further research may reveal that only a nongenomic action explains other functions of calcitriol. Certain effects of this active hormonal form of vitamin D may result from a combination of the two mechanisms, as proposed by Baran et al.46
Age-related changes in vitamin D-driven calcium metabolism
Large numbers of elderly residents in the northern parts of the USA and of Europe have been shown to have depressed circulating levels of 25-OH-D and, in some reports, also low serum concentrations of 1,25(OH)2D. Seasonal differences in 25-OH-D have become well established in these geographic regions,47 8 but l,25(OH)2D insufficiency without symptoms of a fullscale deficiency have only recently been documented.8 9i2K24 Because of the increasing lifespans of so many elderly, especially women, and of residence of large numbers of the elderly in nursing homes or similar facilities that permit little exposure of residents to sunlight, frank deficiency of vitamin D is likely to become more common and even possibly reach epidemic proportions. The public health consequences of osteomalacic bone tissue and increased fracture incidence could be enormous. Thus, some investigators have raised questions about the adequacy of the current allowance of vitamin D, i.e., 200 IU per day, for the elderly.48 50
Human deficiency of vitamin D
Human vitamin D deficiency may result from inadequate dietary intake and/or skin biosynthesis of vitamin D. This relatively prevalent deficiency is characterized by the clinical features and laboratory measurements of blood given in Table 3.
The terms rickets and osteomalacia refer to the histologic and radiologic abnormalities seen in a variety of vitamin D deficiency conditions. (Rickets may also, however, develop from inadequate Ca intake in children without vitamin D deficiency.)51 Rickets describes the disordered growth and mineralization of the growth plate of the long bones.52 The resulting abnormalities, widening of the growth plate, delayed conversion of the growth plate cartilage to bone, and irregularly arranged bone trabeculae, are only seen when the vitamin D deficiency occurs during childhood or adolescence, i.e., before the growth plates close. Osteomalacia describes the abnormalities resulting from delayed and reduced mineralization of mature bone, whether trabecular or cortical. This mineralization defect leads to a widened area of unmineralized bone matrix on the bone surface, called osteoid. Osteomalacia is observed at all ages. Biochemical changes in most cases of rickets and osteomalacia include decreased serum Ca and phosphate levels because of insufficient intestinal absorption of these ions, increased serum parathyroid hormone levels as a result of the reduced serum calcium concentration, and elevated serum alkaline phosphatase concentration. In cases of inadequate supply of vitamin D or in chronic severe liver disease that is associated with fat malabsorption, serum 25-OH-D levels are invariably low, but the levels of l,25-(OH)2D may still be normal, depending on the severity of the condition. Kidney disease, when glomerular filtration rate is less than 30% of normal, will invariably lead to low serum 1,25-(OH)2D levels, while the serum 25-OH-D may be normal.
Table 3 Clinical features and serum changes in human vitamin D deficiency
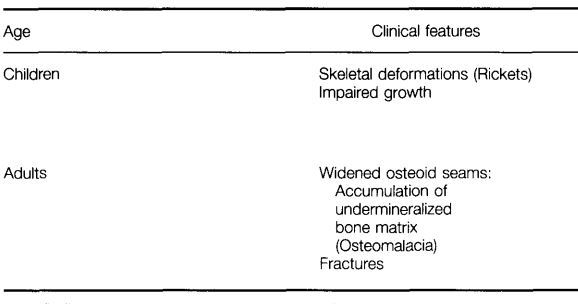
'This finding has not been consistently reported.33
Infants and young children suffer from growth retardation, primarily because of the stunted growth of the long bones, and they may become bowlegged because of the reduced mechanical strength of these bones. Skeletal muscle weakness and increased urinary excretion of amino acids may also occur.
Roentgenographic evidence of rickets includes widened growth plate of the long bones of both arms and legs and reduced bone density. In osteomalacia of the adult, undermineralization of the bones may show up as pseudofractures, which are radiolucent lines on the roentgenograms that mimic stress fractures. However, the pseudofractures are usually bilateral and often involve nonweight-bearing bones, such as the scapula.53
After vitamin D-fortification of foods or the use of cod liver oil was instituted 60 years ago, simple vitamin deficiency rickets became relatively rare in the USA, Canada, and Western Europe. However, a recent increase in the number of such cases in these countries appears to result from either strict vegetarianism, i.e., dietary or religious practices that avoid selection of foods containing vitamin D, or religious dress codes that prevent exposure of the skin to sunlight.54-55 Furthermore, infants who are exclusively breastfed and not given a vitamin D supplement have a high risk of developing rickets,55 particularly black infants of African descent or of other backgrounds whose mothers have a low vitamin D status.56
Causes of rickets or osteomalacia include not only insufficient dietary intake of vitamin D or inadequate sun exposure, but also diseases that may interfere with absorption and biotransformations of vitamin D3 or D2 and conditions leading to decreased tissue responsiveness to the hormone. Patients suffering from malabsorption of fats may have inadequate absorption of vitamin D from the small intestine. Liver disease may interfere with the conversion of vitamin D to 25- OH-D or lead to increased degradation of this metabolite, both of which will lead to decreased circulating levels of 25- OH-D. Patients whose kidneys have been removed or are severely diseased cannot convert 25-OH-D to the hormone, 1,25(OH)2D, and if parathyroid hormone production is inadequate, the vitamin D hormone production will decrease. Finally, three inborn errors of metabolism lead to rickets. Vitamin D-dependent rickets type 1 is caused by a defect in the kidney enzyme, 25-OH-D-1 -hydroxylase, that is responsible for the synthesis of 1,25(OH)2D. In vitamin D-depen- dent rickets type 2, the receptor for 1,25(OH)2D is defective and cannot mediate the genomic action of the hormone. In the third condition, called hypophosphatemic vitamin D-re- sistant rickets, defects in the renal synthesis of 1,25(OH)2D occur as well as in the renal reabsorption of phosphate, leading to low circulating phosphate levels.57
Human toxicity of vitamin D
The human symptoms of vitamin D toxicity (hypervitamin- osis D) are, in general, opposite those of deficiency, as detailed in Table 4.
Toxicity due to vitamin D or its active metabolite may be caused by inadvertent intake of excessive amounts of vitamin D-containing preparations, such as vitamin tablets, or it may occur spontaneously in patients treated with high doses of vitamin D3 or replacement doses of 1,25(OH)2D or an analog of the latter, 1-OH-D, which is converted in the liver to the active hormone. The most consistent and earliest-appearing finding in vitamin D toxicity is hypercalcemia, i.e., serum Ca levels above the normal range of 9.5-11 mg/dL. The serum phosphate level often remains unaltered, but it may also increase acutely, especially if l,25(OH)2D is the offending agent. The cause of the hypercalcemia is most likely increased net Ca influx from bone, and therefore an extension of the normal physiological action of vitamin D; but increased Ca absorption also occurs, and it may contribute to the hypercalcemia.
Table 4 Symptoms of vitamin D toxicity in humans
The increased serum Ca level can lead to a variety of signs and symptoms, such as anorexia,nausea, vomiting, memory loss, confusion, muscle weakness, diarrhea (or constipation), polyuria, and dehydration. The kidneys are particularly sensitive to an elevation of the serum Ca level. First, there is a decrease in the water-concentrating mechanism in the renal tubules, leading to excessive urinary water excretion. Next, Ca salts may form in the renal tubules, giving rise to kidney stones. Finally, if the hypercal- cemic condition persists, Ca salts will be deposited in the renal tissues and cause damage to both the glomeruli and the tubules, with subsequent loss of renal function. Ca salts may be deposited in other soft tissues as well, such as the heart, blood vessels, and skeletal muscles. In general this phenomenon is referred to as metastatic calcification. In long-standing vitamin D toxicity, bone changes characterized by excessive resorption may occur and are referred to as osteitis fibrosa, because part of the bone is replaced by fibrous tissue.4 51
The duration of the patient’s toxic episode is relatively brief if the administered agent is l,25(OH)2D because of the relatively short half-life of the hormone, i.e., 4 to 6 hr. If, however, vitamin D3 or D2 has been administered, the episode could last for weeks or months, because of the higher lipid solubility and storage of these molecules in fat, leading to longer half-lives. When administered at high dose levels, vitamin D3 or D2 will be deposited in body fat and slowly released into the circulation long after dosing is discontinued. Treatment must therefore be continued for a long time to counteract the hypercalcemic response. The treatment includes both drugs to enhance urinary excretion of calcium and drugs to diminish the Ca efflux from bone and absorption of Ca from the intestine.
The sequence of events leading to the hypercalcemia when excessive amounts of vitamin D2 or D3 or 25-OH-D are given is not completely clear. It was previously thought that the high blood levels of 25-OH-D that result from administration of this metabolite or its precursor could result in 25-OH-D binding to and activation of the l,25(OH)2D receptor and, thus, mimicking the action of the hormone itself. However, an alternative explanation is based on the known binding affinities of the different metabolites for the D-binding protein.33 Both vitamin D, and 25-OH-D bind more tightly to the binding protein than l,25(OH)2D does (see Vitamin D binding protein above). Therefore, when the total concentration of 25-OH-D increases manyfold, a certain fraction of the circulating 1,25(OH)2D will be displaced from the binding protein by 25-OH-D and lead to an increase in the free hormone concentration. Thus, the offending agent may be the elevated free hormone concentration even if the total hormone concentration does not increase. Determination of the free hormone fraction has not yet been made in cases of vitamin D toxicity stemming from high doses of 25-OH-D. Consequently, it remains to be seen if the substance interacting with the l,25(OH)2D receptor is the hormone itself or 25- OH-D when excessive doses of vitamin D or 25-OH-D are administered.
Some infants and children may be particularly sensitive to the toxic actions of vitamin D and develop toxicity signs and symptoms after doses as low as 4 to 5 times the recommended daily intake of 400 IU.58 The toxic dose in otherwise healthy adults is probably closer to 30 to 40 times the RDA in the USA.
Summary
Vitamin D, either derived from the diet or skin production, has become an increasingly important molecule in human function because of the extended longevity of human populations in many nations. The actual vitamin D requirements of elderly individuals remain difficult to determine because environmental conditions, especially sunlight exposure, vary so greatly throughout the world. Data are limited on the potential prophylactic benefit of vitamin D or l,25(OH)2D in metabolic bone diseases, such as osteoporosis, in the absence of vitamin D deficiency. Possible roles of vitamin D or its metabolites in the etiology or treatment of diabetes mellitus, cancer, and hypertension have been reviewed recently by Bikle.32 New information on the functional roles of 1,25(OH)2D on target tissues and the mechanism of action of this key molecule continue to emerge.
References
U.S. Department of Agriculture (USDA), Human Nutrition Information Service. (1985). Dietary Levels: Households in the United States, Spring 1977, 1977-78 Nationwide Food Consumption Survey. U.S. Government Printing Office, Washington, DC USA
Meier, D.E., Luckey, M.M., Wallenstein, S., Clemens, T.L., Orwoll, E.S., and Waslien, C.I. (1991). Calcium, vitamin D, and parathyroid hormone status in young white and black women: Association with racial differences in bone mass. .1. Clin. Endocrinol. Metah. 72, 703-710
Holick, M.F., Shao, Q., Liu, W.W., and Chen, T.C. (1992). The vitamin D content of fortified milk and infant formula. New Engl. J. Med. 326, 1178-1181
Jacobus, C.H., Holick, M.F., Shao, Q„ Chen, T.C., Holm, I.A., Ko- lodny, J.M., Fuleihan, G.E.-H., and Seely, E.W. (1992). Hypervitaminosis D associated with drinking milk. New Engl. J. Med. 326, 1173-1177
Subcommittee on Dietary Allowances. (1989). Recommended Dietary Allowances, 10th edition. National Academy Press, Washington, DC USA
Omdahl, J.L., Garry, P.J., Hunsaken, L.A., Hunt, W.C., and Goodwin, J.S. (1982). Nutritional status in a healthy population: Vitamin D. Am. J. Clin. Nutr. 36, 1225-1233
Bouillon, R.A., Auwerx, J.H., Lissens, W.D., and Peleman, W.K. (1987). Vitamin D status in the elderly: seasonal substrate deficiency causes 1,25-dihydroxycholecalciferol deficiency. Am. J. Clin. Nutr. 45,755-763
Egsmose, C., Lund, B., McNair, P, Lund, B., Storm, T., and Sprensen, O. (1987). Low serum levels of 25-hydroxy vitamin D and 1,25-dihydroxy vitamin D in institutionalized old people: influence of solar exposure and vitamin D supplementation. Age Ageing 16, 35-10
Heikinheimo, R.J., Inkovarra, J.A., Harju, E.J., Haavisto, M.V., Kaarela, R.H., Kataja, J.M., Kokko, A.M.-L., Kolho, L.A., and Rajala, S.A. (1992). Annual injection of vitamin D and fractures of aged bones. Calcif. Tissue lnt. 51, 105-110
Dawson-Hughes, B., Dallal, G.E., Krali, E.H., Harris, S., Sokoli, L.J., and Falconer, G. (1991). Effect of vitamin D supplementation on wintertime and overall bone loss in healthy postmenopausal women. Ann. lnt. Med. 115, 505-512
Webb, A.R., Kline, L., and Holick, M.F. (1988). Influence of season and latitude on the cutaneous synthesis of vitamin D,: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D, synthesis in human skin. J. Clin. Endocrinol. Metab. 67,373-378
Stryd, R.P., Gilbertson, T.J., and Brunden, M.N. (1979). A seasonal variation study of 25-hydroxyvitamin D, serum levels in normal humans. J. Clin. Endocrinol. Metab. 48, 771-775
Lawson, D.E.M. (1980). Metabolism of vitamin D. In Vitamin D: Molecular Biology and Clinical Nutrition, (A.E. Norman, ed.), p 93-126, Marcel Dekker, New York, NY USA
Pun, K.K., Wong, F.H.W., Wang, C., Lau, P, Pun, W.K., Chow, S.P., Cheng, C.L., Leong, C.Y., and Young, R.T.T. (1990). Vitamin D status among patients with fractured neck of femur in Hong Kong. Bone 11, 365-368
DeLuca, H.F. (1986). The metabolism and functions of vitamin D. In: Steroid Hormone Resistance: Mechanisms and Clinical Aspects. Adv. Exper. Med. Biol. 196, 361-377
Holick, M.F., MacLaughlin, J.A., Clark, M.B., Holick, S.A., Potts, J.T., Jr., Anderson, R.R., Blank, I.H., Parrish, J.A., and Elias, P. (1980). Photosynthesis of previtamin D, in human skin and the physiologic consequences. Science 210, 203-205
Holick, M.F., MacLaughlin, J.A., and Doppelt, S.H. (1981). Regulation of cutaneous previtamin D, photosynthesis in man: Skin pigment is not an essential regulator. Science 211, 590-593
MacLaughlin, J. and Holick, M.F. (1985). Aging decreases capacity of human skin to produce vitamin D,. J. Clin. Invest. 76, 1536-1538
Reichel, H„ Koeffler, H.P., and Norman, A.W. (1989). The role of the vitamin D endocrine system in health and disease. New Engl. J. Med. 320, 980-991
Sherman, S., Hollis, B„ and Tobin, J. (1990). Vitamin D status and related parameters in a healthy population: The effects of age, sex, and season. J. Clin. Endocrinol. Metab. 71, 405—413
Eastell, R., Yeargey, A., Veiera, N., Cedel, S., Kumar, R., and Riggs, B. (1991). Interrelationship among vitamin D metabolism, true calcium absorption, parathyroid function, and age in women: evidence of an age-related intestinal resistance to 1,25-dihydroxy vitamin D action. J. Bone Miner. Res. 6, 125-132
Mallon, J.P., Matuszewski, D„ and Sheppard, H. (1980). Binding specificity of the rat serum vitamin D transport protein. J. Steroid Biochem. 13, 409^113
Chapuy, M.C., Arlot, M.E., Duboeuf, F., Brun, J., Crouzet, B., Amaud, S., Delmas, P.D., and Meunier, P.J. (1992). Vitamin D, and calcium to prevent hip fractures in elderly women. New Engl. J. Med. ill, 1637-1642
Gallagher, J.C. (1992). Vitamin D metabolism and therapy in elderly subjects. South. Med. J. 85, 43-46 (suppl)
Fujita, T. (1992). Vitamin D in the treatment of osteoporosis. Proc. Soc. Exper. Biol. Med. 199, 394-399
Tilyard, M.W., Spears, G.F.S., Thomsen, J., and Dovey, S. (1992). Treatment of postmenopausal osteoporosis with calcitriol or calcium. New Engl. J. Med. 326, 357-362
Orimo, H., Shiraki, M., Hayashi, T., and Nakamura, T. (1987). Reduced occurrence of vertebral crush fractures in senile osteoporosis treated with la(OH)-vitamin D. Bone Miner. 3, 47-52
Fujita, T. (1990). Studies of osteoporosis in Japan. Metabolism 39, 39-12 (suppl)
Norman, A. (1983). Intestinal calcium absorption: A vitamin D- hormone-mediated adaptive response. Am. J. Clin. Nutr. 51,290-300
Slovik, D.M., Adams, J.S., Neer, R.M., Holick, M.F., and Potts, J.T. (1981). Deficient production of 1,25-dihydroxy vitamin D in elderly osteoporotic patients. New Engl. J. Med. 305, 372-374
Clemens, T.L., Zhou, X.-Y., Myles, M., Endres, D., and Lindsay, R. (1986). Serum vitamin D2 and vitamin D, metabolite concentrations and absorption of vitamin D2 in elderly subjects. J. Clin. Endocrinol. Metab. 63, 656-660
Bikle, D.D. (1992). Clinical counterpoint: Vitamin D: New actions, new analogs, new therapeutic potential. Endocrine Revs. 13, 765-784
Vieth, R. (1990). The mechanisms of vitamin D toxicity. Bone Miner. 11, 267-272
Kawashima, H. and Kurokawa, K. (1986). Metabolism and sites of action of vitamin D in the kidney. Kidney Int. 29, 98-107
Walters, M.R. (1992). Newly identified actions of the vitamin D endocrine system. Endocrine Rev. 13, 719-764
Holick, M.F. (1991). Vitamin D: Cutaneous production and therapeutic effect in psoriasis. In Eighth Workshop on Vitamin D, (A.W. Norman, ed.), p 236, Paris, France (abstr)
Hanchette, C.L. and Schwartz, G.G. (1992). Geographic patterns of prostate cancer mortality: Evidence for a protective effect of ultraviolet radiation. Cancer 70, 2861-2869
Pike, J.W. (1991). Vitamin D, receptors: Structure and function in transcription. Ann. Rev. Nutr. 11, 189-216
Halloran, B.P. and DeLuca, H.F. (1981). Appearance of the intestinal cytosolic receptor for 1,25-dihydroxyvitamin D3 during neonatal development in the rat. J. Biol. Chem. 256, 7338-7342
Wasserman, R.H. and Fullmer, C.S. (1983). Calcium transport proteins, calcium absorption, and vitamin D. Ann. Rev. Physiol. 45, 375-390 '
Darwish, H.M. and DeLuca, H.F. (1992). Identification of a 1,25- dihydroxyvitamin D3-response element in the 5'-flanking region of the rat calbindin D-9k gene. Proc. Natl. Acad. Sci. (USA) 89, 603-607
Nemere, I. and Norman, A.W. (1990). Transcaltachia, vesicular calcium transport, and microtubule-associated calbindin-D28K: Emerging views of 1,25-dihydroxyvitamin D,-mediated intestinal calcium absorption. Miner. Electrolyte Metab. 16, 109-114
Wali, R.K., Baum, C.L., Sitrin, M.D., and Brasitus, T.A. (1990).
1,25(OH), vitamin D3 stimulates membrane phosphoinositide turnover, activates protein kinase C, and increases cytolsolic calcium in rat colonic epithelium. J. Clin. Invest. 85, 1296-1303
Lieberherr, M., Grosse, B„ Duchambon, R, and Drueke, T. (1989). A functional cell surface type receptor is required for the early action of 1,25-dihydroxyvitamin D3 on the phosphoinositide metabolism in rat enterocytes. J. Biol. Chem. 264, 20403-20406
Norman, A.W., Nemere, I„ Zhou, L.-X., Bishop, J.E., Lowe, K.E., Maiyar, A.C., Collins, E.D., Taoka, T„ Sergeev, I„ and Farach- Carson, M.C. (1992). 1,25(OH)2-vitamin D3, a steroid hormone that produces biological effects via both genomic and nongenomic pathways. J. Steroid Biochem. Molec. Biol. 41, 231-240
Baran, D.T., Sorensen, A.M., Shalhoub, V., Owen, T., Stein, G., and Lian, J. (1992). The rapid nongenomic actions of la,25-dihydroxy- vitamin D, modulate the hormone-induced increments in osteocalcin gene transcription in osteoblast-like cells. J. Cell. Biochem. 50, 124-129
Tjellesen, L. and Christiansen, C. (1983). Vitamin D metabolites in normal subjects during one year. A longitudinal study. Scand. J. Clin. Lab. Invest. 43, 85-89
Parfitt, A.M., Gallagher, J.C., Heaney, R.P., Johnston, C.C., Jr., Neer, R., and Whedon, G.D. (1982). Vitamin D and bone health in the elderly. Am. J. Clin. Nutr. 36, 1014—1031
Webb, A.R., Pilbeam, C., Hanafin, N„ and Holick, M.F. (1990). An evaluation of the relative contributions of exposure to sunlight and of diet to the circulating concentrations of 25-hydroxyvitamin D in an elderly nursing home population in Boston. Am. J. Clin. Nutr. 51, 1075-1081
Gloth, F., Ill, Tobin, J., Sherman, S., and Hollis, B. (1991). Is the Recommended Daily Allowance for vitamin D too low for the homebound elderly? J. Am. Geriatr. Soc. 39, 137-141
Okonofua, F., Gill, D.S., Alabi, Z.O., Thomas, M., Bell, J.L., and Dandona, P. (1991). Rickets in Nigerian children: A consequence of calcium malnutrition. Metabolism 40, 209-213
David, L. (1991). Common vitamin D-deficiency rickets. In Rickets, (F.H. Glorieux, ed.), p 107-122, Raven Press, New York, NY USA
Pitt, M.J. (1991). Rickets and osteomalacia are still around. Radiol. Clin. N. Am. 29, 97-119
Dagnelie, PC., Vergote, F.J.V.R.A., vanStaveren, W.A., vandenBerg, H„ Dingjan, P.G., and Hautvast, J.G.A.J. (1990). High prevalence of rickets in infants on macrobiotic diets. Am. J. Clin. Nutr. 51, 202-208
Markestad, T. and Elzouki, A.Y. (1991). Vitamin D-deficiency rickets in Northern Europe and Libya. In Rickets, (F.H. Glorieux, ed.), p 203-213, Raven Press, New York, NY USA
Chang, Y.T., Germain-Lee, E.L., Doran, T.F., Migeon, C.J., Levine, M.A., and Berkovitz, G.D. (1992). Hypocalcemia in nonwhite breast-fed infants. Clin. Pediatr. 31, 695-698
Glorieux, F.H.. (1991). Preface. In Rickets. (F.H. Glorieux, ed), Raven Press, New York, NY USA
Jeans, PC. and Steams, G. (1938). The effect of vitamin D on linear growth in infancy. II. The effect of intakes above 1,800 USP units daily. J. Pediatr. 13, 730-740
Chesney, R., Rosen, J„ Hamstra, A., and DeLuca, H. (1980). Serum 1,25-dihydroxyvitamin D levels in normal children and in vitamin D disorders. Am. J. Dis. Child 134, 135-139
Rosen, J. and Chesney, R. (1983). Circulating calcitriol concentrations in health and disease. J. Pediatr. 103, 1-17