Increased Vitamin D increases Phosphate as well as Calcium
Increased intestinal phosphate absorption, an often-overlooked effect of vitamin D
R. Todd Alexander1,2,3 todd2@ualberta.ca
1 Department of Physiology University of Alberta, Edmonton, T6G 1C9, Canada
2 Pediatrics, University of Alberta, Edmonton, T6G 1C9, Canada
3 Membrane Protein Disease Research Group, University of Alberta, Edmonton, T6G 1C9 Canada
 * ATP is the source of energy in the body = Adenosine Tri phosphate = C10H16N5O13 P3
* each ATP molecule is recycled some 1000 to 1500 times per day.
* A body creates 160 kilograms of ATP daily
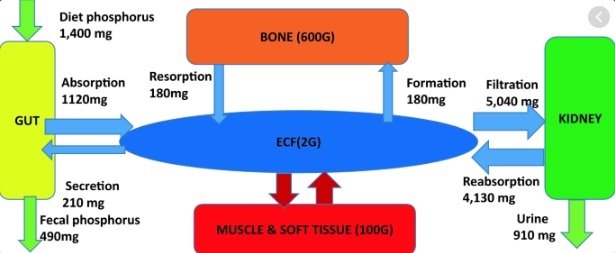
* Phosphorus in Your Diet You need phosphorus to:
* keep your bones strong and healthy
* help make energy
* move your muscles
* build strong teeth
* manage how your body stores and uses energy
* reduce muscle pain after exercise
* filter out waste in your kidneys
* grow, maintain, and repair tissue and cells
* produce DNA and RNA — the body’s genetic building blocks
* balance and use vitamins such as vitamins B and D, as well as other minerals like iodine, magnesium, and zinc
* maintain a regular heartbeat
* facilitate nerve conduction
* ATP is the source of energy in the body = Adenosine Tri phosphate = C10H16N5O13 P3
* each ATP molecule is recycled some 1000 to 1500 times per day.
* A body creates 160 kilograms of ATP daily
* Phosphorus in Your Diet You need phosphorus to:
* keep your bones strong and healthy
* help make energy
* move your muscles
* build strong teeth
* manage how your body stores and uses energy
* reduce muscle pain after exercise
* filter out waste in your kidneys
* grow, maintain, and repair tissue and cells
* produce DNA and RNA — the body’s genetic building blocks
* balance and use vitamins such as vitamins B and D, as well as other minerals like iodine, magnesium, and zinc
* maintain a regular heartbeat
* facilitate nerve conduction
📄 Download the PDF from sci-hub via VitaminDWiki
The oldest known and most important function of the active form of vitamin D, calcitriol, is to increase intestinal calcium absorption, and thus serum calcium concentration. The mechanism of action whereby calcitriol enhances intestinal calcium absorption is through increasing active transcellular and likely also passive paracellular calcium absorption (reviewed in ⑴).The former effect is primarily mediated by increasing transcription of the apical calcium channel Trpv6, which enhances intestinal epithelial calcium entry and thus calbindin-D9K expression. Calcitriol likely enhances paracellular calcium absorption by increasing the transcription of claudin-2 and claudin-12, which contribute intestinal paracellular calcium pores. However, when luminal calcium levels are lower than plasma, having more intestinal calcium permeability may be counterproductive, and enhance calcium secretion and thus loss in feces (2). Clearly there are further questions with respect to how intestinal calcium absorption is increased in response to calcitriol.
The second major effect of calcitriol on intestinal mineral transport is to increase phosphate absorption. However, much less is known about how calcitriol mediates this. Increased sodium phosphate cotransporter 2b (NaPi2b, also known as NaPi-IIb, NPTIIb, and NPT2b) protein expression has been implicated, but if other apical phosphate transporters contribute, and if paracellular phosphate absorption is enhanced by calcitriol is unknown. Paracellular phosphate absorption appears, like calcium, to be the predominant pathway by which these minerals are absorbed from the gut (3). Yet the molecular details of the paracellular pathway are completely unknown. Moreover, plasma phosphate levels, which are in part regulated by intestinal absorption, associate with all-cause mortality, even in the “normal” range, making this area of enquiry all the more important.
To delineate the mechanism whereby calcitriol increases intestinal phosphate absorption Hernando et al administered calcitriol to wild-type and Slc34a2 knockout (KO) mice (N.B. Slc34a2 encodes the sodium-phosphate cotransporter, NaPi2b) and examined intestinal phosphate absorption ⑷.They found no effect of calcitriol administration on phosphate absorption in the ileum. However, Using chamber studies performed on the jejunum ex vivo demonstrated increased radioactive phosphate flux in wild-type, but not Slc34a2 KO mice, in the presence of low apical phosphate concentration (i.e. under conditions strongly favouring transcellular phosphate absorption). Consistent with the functional studies, NaPi2b protein but not Slc34a2 mRNA, was increased in the jejunum of wild-type mice in response to calcitriol. These results strongly support NaPi2b mediated apical phosphate uptake being the only transcellular pathway responsible for increasing small intestinal phosphate absorption in response to calcitriol administration.
The authors also interrogated the effect of calcitriol on the paracellular pathway. They did so by examining phosphate flux across jejunal and ileal segments ex vivo in Ussing chambers in the presence of large phosphate concentration gradients (70 mM, i.e. conditions strongly favouring paracellular diffusive flux). They did not observe a difference in paracellular phosphate absorption in either genotype in response to calcitriol. Although, interestingly, the Slc34a2 KO mice displayed reduced paracellular phosphate flux across the ileum by this method. To confirm that calcitriol had no effect on the paracellular pathway they determined paracellular phosphate permeability of the jejunum and ileum ex vivo in Ussing chambers by measuring phosphate to chloride bionic diffusion potentials. This failed to detect a difference in phosphate permeability in either genotype, before or after treatment with calcitriol. Thus, in the small intestine at least, it appears that calcitriol does not enhance intestinal phosphate absorption by increasing paracellular phosphate absorption.
This work contributes much to our understanding of intestinal phosphate absorption but also raises some questions. Consistent with the data reported here, others have seen increased NaPi2b protein but not mRNA expression in response to calcitriol treatment, begging the question by what post-translational mechanism does this occur? Hernando et al report significant expression of Slc30a1 and Slc30a2 mRNA in the colon, although expression was not altered in the Slc34a2 KO animals nor by calcitriol (4). Protein expression was not assessed. However, this raises the possibility that there could be an effect of calcitriol on colonic phosphate absorption? Further, why was less paracellular phosphate flux observed in response to a large chemical gradient, across the ileum, but not the jejunum, of Slc34a2 KO mice? This is particularly intriguing as phosphate permeability was not lower in the ileum of the KO mice, and both the lumen negative potential difference and concentration difference across the tissue were the same in both genotypes. Moreover, why was this difference not observed in a previous study from this group (5)? Ultimately, this work convincingly demonstrates that NaPi2b is the sole small intestinal regulator of increased phosphate absorption in response to calcitriol and that there is no effect on paracellular phosphate absorption. It also serves to highlight a central question in the field, what is the molecular identity of the paracellular phosphate pore in the lntestine?