Less radiation therapy needed if have more vitamin D (lung cancer in lab)
Vitamin D improves the sensitization of lung cancer to radiotherapy
Int J Clin Exp Med 2017;10(3):4921-4928 www.iicem.com /ISSN:1940-5901/IJCEM0033582
Zongye Wang1, Junli Liu2, Shaoyan Si2, Bingxin Xu2, Ying Wang2, Ye Ren1, Zhuojie Dai1, Di Cui1, Xiaoming Su1, Shujun Song2
1Department of Therapeutic Radiology and Oncology, 306 Hospital of PLA, Beijing, P.R. China; 2Center for Special Medicine and Experimental Research, 306 Hospital of PLA, Beijing, P.R. China
Received January 10, 2016; Accepted October 29, 2016; Epub March 15, 2017; Published March 30, 2017
 1. Figure 4 shows
* Radiation by itself increases lung cancer cell death.by 6
* Radiation & active vitamin D increases lung cancer cell death by 11 ( almost double)
1. Figure 4 shows
* Radiation by itself increases lung cancer cell death.by 6
* Radiation & active vitamin D increases lung cancer cell death by 11 ( almost double)
 1. Note: VitaminDWiki is exploring the use of inhaled Vitamin D to directly treat respiratory problems.
* In 2016 lung tissue (in mice) was found to directly activate inhaled vitamin D
* Perhaps inhaled vitamin D could potentiate lung cancer radiation therapy and/or chemotherapy
* Thus less conventional cancer therapy might be needed with inhaled vitamin D
* see Inhaled vitamin D help lungs - many studies
* You can make your own inhaled vitamin D with products avaiable on Amazon
1. See also VitaminDWiki
* * Cancer - After diagnosis category listing has items along with related searches**
* Vitamin D and breast cancer radiation - with observations on vitamin D and radiation therapy
* 3X less pelvic inflammation from cancer radiation if high level of vitamin D – Feb 2015
* Both Vitamin D and Vitamin C fight cancers and aid anticancer drugs – May 2018
* Radiation therapy for cancer should be helped by Vitamin D – April 2021
* Chemotherapy and vitamin D - many studies
*
1. Note: VitaminDWiki is exploring the use of inhaled Vitamin D to directly treat respiratory problems.
* In 2016 lung tissue (in mice) was found to directly activate inhaled vitamin D
* Perhaps inhaled vitamin D could potentiate lung cancer radiation therapy and/or chemotherapy
* Thus less conventional cancer therapy might be needed with inhaled vitamin D
* see Inhaled vitamin D help lungs - many studies
* You can make your own inhaled vitamin D with products avaiable on Amazon
1. See also VitaminDWiki
* * Cancer - After diagnosis category listing has items along with related searches**
* Vitamin D and breast cancer radiation - with observations on vitamin D and radiation therapy
* 3X less pelvic inflammation from cancer radiation if high level of vitamin D – Feb 2015
* Both Vitamin D and Vitamin C fight cancers and aid anticancer drugs – May 2018
* Radiation therapy for cancer should be helped by Vitamin D – April 2021
* Chemotherapy and vitamin D - many studies
* 
📄 Download the PDF from VitaminDWiki
Abstract: Lung cancer is the leading cause of cancer related death worldwide. Radioresistance is an obstacle in treating lung cancer. The efficacy of treatments may be improved by increasing the sensitivity of cancer cells to radiation therapy. Vitamin D has been reported to have radiosensitizing effects in cancers of the breast and the prostate .
The objective of this study is to explore the effects of vitamin D on radiosensitization of human lung cancer cells. Human non-small cell lung cancer cell line A549 cells were treated with 1,25(OH)2D3 before irradiation with X-rays. The effect of 1,25(OH)2D3 on cell-survival following irradiation was evaluated by colony-forming assay. DNA double-strand breaks were detected by immunostaining for yH2AX foci. Cell cycle distributions were investigated using flow cytometry. Apoptotic cells were identified on the basis of nuclear morphology. Results: 1,25(OH)2D3 inhibited lung cancer cell proliferation in a dose-dependent manner. A549 cells were radiosensitized by 1,25(OH)2D3. Flow cytometric analysis for DNA content indicated that 1,25(OH)2D3 increased radiation-induced G2/M arrest. Apoptotic analysis demonstrated that 1,25(OH)2D3 enhanced the cell apoptosis induced by irradiation. Conclusion: These findings suggest that 1,25(OH)2D3 caused radiosensitization of A549 cells are associated with its enhancement effects on apoptosis and G2/M arrest induced by radiation.!!
Introduction
Lung cancer is one of the most frequently diagnosed cancers and the leading cause of cancer related death worldwide. Its rates have been steadily increasing in recent years. Non-small cell lung cancer (NSCLC) is accounted for more than 80% of the total lung cancers [1]. Currently, the epidemiologic survey displayed that the 5 year survival rate of patients in the Stage I NSCLC was about 55%~72%. Unfortunately, some unresectable cases such as patients with NSCLC of stage II, III B and IV only had less than 5% the 5 year survival rate [2]. Radiotherapy provides a clear survival advantage for patients with NSCLC and remains to be an effective measure in achieving the local control of the disease. However, the development of radioresistance frequently influences the efficacy of radiotherapy and thus becomes an obstacle of this technique. Therefore, it would be desirable to develop agents that could potentiate the effects of radiation. Ideally, such agents would be non-toxic and enhance the killing effect of radiation on cancer cells.
Vitamin D is an essential fat-soluble vitamin. 1a, 25-dihydroxy-cholecalciferol (1,25(OH)2D3), also known as calcitriol, is an active form of vitamin D. It have been confirmed that vitamin D plays an important role in maintaining the calcium homeostasis and bone mineralization. Recent years, 1,25(OH)2D3, as an important bone regulator, has been suggested to have multiple functions, including anticancer effect
. The most of the anti-cancer actions of vitamin D have been studied in cancers of breast, prostate, and colon. Studies have provided evidence that circulating 25(OH)D levels are inversely associated with the incidence of colorectal cancer [4], prostate cancer risk [5] and breast cancer [6]. There are also investigations about vitamin D and lung cancers. Results of a meta-analysis showed an inverse association between serum vitamin D and lung cancer risk [7]. Evidences in the studies of patients with early stage NSCLC in US and Norwegian showed that high level of vitamin D may prolong survival time [8, 9]. Studies showed Vitamin D intake was associated with a lower lung cancer risk in never-smoking, postmenopausal women
. An in vitro study showed that vitamin D inhibited the growth of lung cancer cell lines
. Findings of other studies suggest that vitamin D could inhibit the growth and metastasis of lung cancer in mouse models [12, 13]. These findings support the potential of vitamin D as a therapeutic agent for lung cancer. As its antitumor capacity, vitamin D might be also considered as a chemosensitizer or radiosensitizer. It has been found that 1,25(OH)2D3 or its synthetic analogues increased the susceptibility of cancer cells to the cytotoxic action of tumor necrosis factor [14], adriamycin and paclitaxe [15]. Vitamin D3 or its analog has also been shown to enhance the responses of breast cancer cells [16, 17] and prostate cancer cells [18] to radiation. Vitamin D enhances cancer cells response to radiation by decreasing cell viability, inducing cell apoptosis and cell cycle arrest [16, 19]. However, its radiosensitizing effects are not well studied in lung cancer. The aim of this study is to explore the potential of vitamin D as a radiosensitizer in lung cancer.
Materials and methods
Materials and cell cultures
The NSCLC cell line (A549) were obtained from ATCC (Manassas, VA, USA) and maintained at 37°C in humidified incubator containing 5% CO2 in RPMI-1640 (Gibco Invitrogen, Burlington, ON, USA) containing 10% fetal calf serum (Gibco Invitrogen, Burlington, ON, USA) with supplements. All other chemicals and reagents were purchased from Sigma Chemical Co. (St. Louis, MO, USA) unless otherwise stated. 1,25(OH)2D3 (0.1 mol/L) was dissolved in absolute ethanol and further diluted in cell culture medium as required concentrations. Control cells were always treated with the corresponding concentrations of ethanol.
Cell proliferation assessment by MTT assay
Cells were seeded at 3,000 per well in 96-well plates, allowed to attach overnight and cells were treated with different concentrations (50- 1000 nmol/L) of 1,25(OH)2D3. Medium containing ethanol served as the controls. After 72 hour incubation, cell proliferation was assessed using 3-(4,5 dimethylthiazol-2-yl)-2,5-diphenyl- tetrazolium bromide (MTT) based on colorimetric assay as previously described [20]. The absorbance was measured at 490 nm using a microplate reader (BIO RAD, Hercules, CA, USA). Cellular proliferation was expressed as a percentage with vehicle-treated cells.
Irradiations
Radiation was administered with 6 MV of x-rays from a linear accelerator (Elekta Axesse, Elekta Medical Equipment Co. Ltd, Sweden) with graded doses (0, 2, 4, 6 and 8 Gy).
Clonogenic analysis of cell survival
Cells were seeded in 6 well plates at a density of 200 cells per well. After attachment, cells were treated with 1,25(OH)2D3 (100 nmol/L) or vehicle for 24 hours. The cells were then unirradiated (control) or irradiated for total doses ranging from 0 to 8 Gy. Media containing 1,25(OH)2D3 was exchanged for fresh media. Cells were subsequently cultured under standard cell culture conditions for 8 days, with media were changed every third day. Colonies were stained with Giemsa solution, and colonies containing at least 50 cells in size were counted. The surviving fraction (SF) was calculated based on the plating efficiency of nonir- radiated control cultures treated with the vehicle. The survival curve was plotted using X-Y log scatter (Graph Prism 5.0). Based on the clonogenic survival assay, the dose enhancement ratio (DER) was calculated as the ratio of radiation doses required to give the same biological effect in the absence or the presence of 1,25(OH)2D3. The DER for 10% survival was determined. A DER greater than one is indicated radiosensitization.
Y-H2AX immunofluorescence staining of cells
Cells were cultured onto cover slips in 24-well plates and incubated overnight. And then cells were treated with 1,25(OH)2D3 (100 nmol/L) for 24 hours, then irradiated (0 or 4 Gy). At 1 hour and 24 hours post radiation, cells were fixed with 4% formaldehyde, the cells were rinsed in PBS and permeabilized with 0.2% Triton X100 in PBS for 20 minutes at room temperature and blocked in PBS containing 5% normal goat serum for 30 min. Slips were incubated with Alexa Fluor 488-conjugated rabbit-anti-y-his- tone-H2AX antibody (Cell Signaling Technology, Danvers, MA, USA) at a dilution of 1:500 in 5% bovine serum albumin at 4°C overnight. After washing with PBS, Slips were rinsed and mounted in Gelvatol (25% polyvinyl alcohol, 30% glycerol, and 0.1% sodium azide) containing 4',6-diamidino-2-phenylindole (DAPI, 1 |Jg/ml). Cover slips were then mounted on glass slides, and cells were analyzed with an Olympus BX51 fluorescent microscope (Olympus Corporation, Tokyo, Japan) using 40x objective. The images were captured using a digital camera (Olympus DP72). Cells were classified positive (containing radiation-induced H2AX foci) when more than five foci were detected. Total and Y-H2AX- positive cells were counted in at least five representative fields per cover slip using a 40x objective.
Cell cycle analysis
Cells were seeded in 6 well plates at 1.5*105 cells per well and incubated overnight. Cells were then treated with 1,25(OH)2D3 at 0 or 100 nmol/L for 24 hours. Cells were irradiated with 0 or 4 Gy of X-rays and incubated in a medium containing the drug for appropriate time (0, 6, 12, 24, 48 h). They were then fixed with 70% ethanol, treated with RNase A and incubated in propidium iodide solution after washing. Cell cycle distribution was analyzed using BD FACSCalibur (Becton, Dickinson and Company, NJ, USA).
Apoptotic assay
The apoptosis of cells were analyzed as previously described [21]. Cells were cultured onto cover slips in 24-well plates and incubated overnight. And then cells were treated with 1,25(OH)2D3 (100 nmol/L) for 24 hours, then irradiated with 0 or 4 Gy of X rays. Three days after radiation, cells were fixed with 4% formaldehyde and rinsed in PBS. The cells were per- meabilized with 0.2% Triton X100 in PBS for 20 minutes. The cover slips were rinsed and mounted in Gelvatol containing DAPI. Fluorescent images were captured with a digital camera linked to a fluorescent microscope. Total and apoptotic nuclei were counted at least in five representative fields per cover slip using a 40x objective. Apoptotic cells were identified on the basis of morphology (nuclear condensation and fragmentation). Values for apoptotic cells were expressed as the percentage of total cells.
Statistical methods
Statistical analysis was performed using Statistical Product and Service Solutions (SPSS) software version 16.0 (SPSS Inc, Chicago, IL, USA). Data were presented as mean ± SD. The differences between groups were tested for significance using the one-way analysis of variance or student t test P < 0.05 was considered statistically significant.
Results
1,25(OH)2D3 inhibits proliferation of A549 cells
The proliferation assay was performed to evaluate the effects of 1,25(OH)2D3 on A549 Cell growth inhibition. It was found that 1,25(OH)2D3 dose dependently inhibited A549 cell growth. Treatment with 200 nmol/L or 500 nmol/L of 1,25(OH)2D3 showed approximately 40% or 90% inhibition of A549 cells growth (Figure 1). Concentrations greater than 500 nmol/L did not result in further growth inhibition. 1,25(OH)2D3 had no inhibitory effects on cell proliferation when concentrations were at or lower than 100 nmol/L (Figure 1). To test its potential as a radiosensitizer, 1,25(OH)2D3 at 100 nmol/L was used in the subsequent experiments.
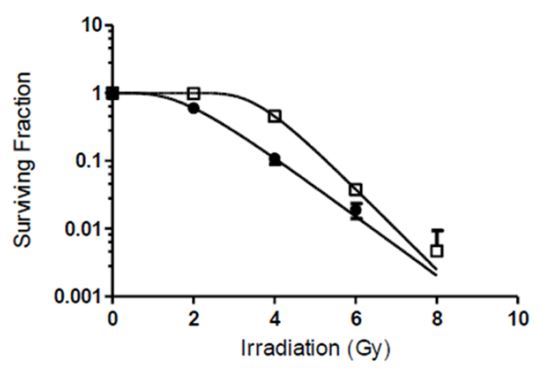
Vitamin D sensitizes A549 cells to radiation
To determine whether vitamin D could radio- sensitize A549 cells, cells were exposed to 0 or 100 nmol/L 1,25(OH)2D3 for 24 hour, and then irradiated with 0, 2, 4, 6 or 8 Gy of X-rays. As shown in Figure 2, A549 cells treated only with 100 nmol/L 1,25(OH)2D showed no effect on cell survival. However, A549 cells pretreated with 1,25(OH)2D3 plus X-ray irradiation exhibited significantly lower clonogenic survival fractions than cells treated with radiation alone. Survival fraction at 4 Gy was reduced from 45.5% in the irradiated- only controls to 10.8% in 1,25(OH)2D3 treated and irradiated cells. The DER for 10% cell survival was 1.33 (Figure 2). 1,25(OH)2D3 treatment significantly sensitized the cells to irradiation.
1,25(OH)2D3 does not enhance the radiation-induced DNA damage or inhibit the DNA damage repair
Y-H2AX has been identified as a marker of DNA double-strand break. In order to determine the effects of 1,25(OH)2D3 on DNA damage, immunofluo- renscent staining using the anti-Y-H2AX antibodies was conducted. Y-H2AX foci could be clearly distinguished 1 hour post 4 Gy radiation. Y-H2AX positive cells were shown to be reduced dramatically at 24 hour time point. The percentages of Y-H2AX positive cells were no difference between cell treated with a combination of 1,25(OH)2D3 and radiation and treated with radiation alone at either 1 hour or 24 hour time point. These results implicate that the DNA damage or damage repair is not influenced by the presence of vitamin D.
1,25(OH)2D3 pretreatment enhances radiation- induced G2/M arrest
To test the radiosensitization nature of 1,25- (OH)2D3, cell cycle was evaluated by flow cytometry. As shown in Figure 3, radiation induced cell cycle arrest in G2/M phase at 6, 12 and 24 hour time points and the altered cell cycle recovered 48 hours after irradiation. However, cells pretreated with 1,25(OH)2D3 showed more cells arrested in G2/M phase at 48 time point compared with cells treated with radiation alone (13.7 ± 1.0% vs 10.3 ± 0.3%, P < 0.05; Figure 3). 1,25(OH)2D3 alone had no effect on cell cycle phase distribution.
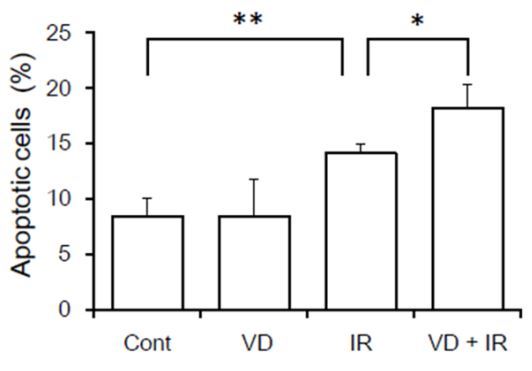
The treatment withl, 25(OH)2D3 increases radiation-induced A549 cell apoptosis
Apoptosis assay performed after 72 hour irradiation and demonstrated that 4 Gy irradiated cells pretreated with VD had more apoptotic cells (18.2 ± 2.2%) than cells treated with irradiation alone (14.1 ± 0.9%, P < 0.05; Figure 4). 1,25(OH)2D3 (100 nmol/L) alone had no effects on A549 cell apoptosis.
Discussion
Recent years, vitamin D has been suggested to have multiple functions, including anticancer effect [22, 23]. Vitamin D may prevent some type of cancers such as breast cancer, colon cancer and prostatic cancer. Inhibition of cancer cell growth is suggested to be one of the mechanisms of its anticancer effects [23]. In the present study we demonstrate that vitamin D dose-dependently inhibit the growth of A549 cells. These findings add to the evidence that vitamin D may have anti-cancer effects.
Given the ability of 1,25(OH)2D3 to inhibits the growth of some type of cancer cells, we sought to investigate its potential as a radiosensitizer in lung cancer cells. Radiosensitizing effects of 1,25(OH)2D3 have been reported in breast cancer cells [16, 17] and prostate cancer cells [18]. In the present study using clonogenic assays, we assessed the effects of vitamin D on lung cancer cell survival. Our results showed that vitamin D pretreatment followed by radiation exposure dramatically reduced colony formation compared to radiation alone. The present results indicate that 1,25(OH)2D3 pretreatment enhance the cell killing and showed a significant radiosensitizing effect on the lung cancer cell line.
The mechanism of vitamin D-induced radiosensitization remains largely unknown at the cellular level. Increasing radiation-induced DNA damage and decreasing DNA damage repair have been demonstrated to play roles in tumor radiosensitivity. Y-H2AX, as a marker of DNA double-strand break, has shown to be positively associated with tumor radiosensitivity [24]. In the present study, the DNA damage was assessed by measuring the expression levels of Y-H2AX in response to radiation and radiation combined with 1,25(OH)2D3. Our results demonstrate that 1,25(OH)2D3 fails to augment DNA damage induced by radiation or inhibit DNA damage repair in A549 cells, which is consistent with findings in a previous study [25]. These findings suggest that vitamin D promoted lung cancer cell killing induced by radiation may not correlate to the alteration of DNA damage and DNA damage repair. Similar finding was also observed in breast cancer cells [26, 27].
The use of different agents to arrest cells in the radiosensitive phases of the cell cycle is a strategy for radiosensitization. Cells are most sensitive to the effects of radiation in G2/M phase and most resistant in G0/G1 [28]. Studies have shown that the radiosensitizing effect of some anticancer drugs is due to cell cycle alteration, such as Paclitaxel and Cucurbitacin B. These drugs Potentiate radiosensitivity by mainly blocking cells in G2/M phase of the cell cycle [29, 30]. To investigate the mechanism by which vitamin D enhance lung cancer cell sensitivity to radiation, we examined the effects of combined treatment or radiation alone on cell cycle regulation by flow cytometry. Our results demonstrated a slight but statistically significant increase in the proportion of cells in G2/M phase for cells receiving combined treatment of radiation and 1,25(OH)2D3 than cells receiving radiation alone. The results suggest that the enhanced radiation effects by vitamin D seem to be associated with cell cycle arrest. Therefore, cell cycle regulation may be one of the mechanisms of the effects of vitamin D on the radiosensitization. However, in the present study the cell cycle regulating effect of 1,25(OH)2D3 is modest. Therefore, cell cycle redistribution does not appear to play a major role in vitamin D-induced radiosensitization. Other mechanisms, such as apoptosis, might contribute to the increased cell killing observed in the clonogenic assays.
Apoptosis is a programmed cell death. It is important defense mechanisms of the system to either repair the damage or eliminate the defective cells (containing damages) [31]. Apoptosis has previously been regarded as a potential mechanism of radiosensitization. Different studies have reported the role of apoptosis in radiosensitization [32, 33]. Apoptosis enhancement in vitamin D induced radiosensitization has been observed in prostate cancer cells [18] and breast cancer cells [16]. It has been suggested that pretreatment with 1,25(OH)2D3 lowers the apoptotic threshold of cancer cells to the subsequent stress of radiation [18]. Increased apoptosis of cancer cells may result in reoxygenation for hypoxic cancer cells, and thus contribute to cytotoxicity of subsequent radiation treatments [34, 35]. In the present study we showed that pretreatment with 1,25(OH)2D3 could significantly enhance the radiation-induced apoptosis in A549 cells, which could be also one of the mechanisms underlying the sensitization of lung cancer cells to the effects of radiation. However, a previous study showed that apoptosis enhancement was not observed in A549 cells co-treated with 1,25(OH)2D3 and radiation [25]. The inconsistent results may attribute to different experimental conditions. In their experiment, they used 137Cs gamma rays, but X-rays were used in our experiments. The duration of total exposure time to 1,25(OH)2D3 was also different in these two studies. The results in our study suggest that vitamin D may increase the efficacy of irradiation-induced lung cancer cell death. However, further investigations and characterization are need for evaluating radiosensitizing effects of vitamin D.
In conclusion, vitamin D radiosensitized A549 cells by significantly decreasing the cell colony formation ability. Enhanced G2/M cell cycle arrest and apoptosis could be possible mechanisms underlying the effects of vitamin D on radiosensitization. The finding implicates that vitamin D may be a useful radiation sensitizer in human lung cancer cells.
Acknowledgements
This work was supported by the Medical Research Grant of 306 Hospital of PLA. The authors wish to thank Yaya Qin, Xiaoqing Tan and the staff in the Department of Therapeutic Radiology and Oncology in 306 Hospital of PLA for their technical assistance.
Disclosure of conflict of interest: None.
Address correspondence to: Dr. Shujun Song, Center for Special Medicine and Experimental Research,306 Hospital of PLA, 9 North Anxiang Road,
Chaoyang District, Beijing 100101, P.R. China. Tel:86-10-66356729-2344; Fax: +86-10-66356306;E-mail: shuj80@126.com
References
Gandara D, Narayan S, Lara PN Jr, Goldberg Z, Davies A, Lau DH, Mack P, Gumerlock P, Vijayakumar S. Integration of novel therapeutics into combined modality therapy of locally advanced non-small cell lung cancer. Clin Cancer Res 2005; 11: 5057s-5062s.
Reungwetwattana T, Eadens MJ, Molina JR. Chemotherapy for non-small-cell lung carcinoma: from a blanket approach to individual therapy. Semin Respir Crit Care Med 2011; 32: 7893.
Feldman D, Krishnan AV, Swami S, Giovannucci E, Feldman BJ. The role of vitamin D in reducing cancer risk and progression. Nat Rev Cancer 2014; 14: 342-357.
Garland CF, Garland FC, Gorham ED, Lipkin M, Newmark H, Mohr SB, Holick MF. The role of vitamin D in cancer prevention. Am J Public Health 2006; 96: 252-261.
Corder EH, Friedman GD, Vogelman JH, Orentreich N. Seasonal variation in vitamin D, vitamin D-binding protein, and dehydroepian- drosterone: risk of prostate cancer in black and white men. Cancer Epidemiol Biomarkers Prev 1995; 4: 655-659.
Garland CF, Gorham ED, Mohr SB, Grant WB, Giovannucci EL, Lipkin M, Newmark H, Holick MF, Garland FC. Vitamin D and prevention of breast cancer: pooled analysis. J Steroid Bio- chem Mol Biol 2007; 103: 708-711.
Zhang L, Wang S, Che X, Li X. Vitamin D and lung cancer risk: a comprehensive review and meta-analysis. Cell Physiol Biochem 2015; 36: 299-305.
Tretli S, Schwartz GG, Torjesen PA, Robsahm TE. Serum levels of 25-hydroxyvitamin D and survival in Norwegian patients with cancer of breast, colon, lung, and lymphoma: a population-based study. Cancer Causes Control 2012; 23: 363-370.
Zhou W, Heist RS, Liu G, Asomaning K, Neuberg DS, Hollis BW, Wain JC, Lynch TJ, Giovannucci E, Su L, Christiani DC. Circulating 25-hydroxyvitamin D levels predict survival in early-stage non-small-cell lung cancer patients. J Clin Oncol 2007; 25: 479-485.
Cheng TY, Lacroix AZ, Beresford SA, Goodman GE, Thornquist MD, Zheng Y, Chlebowski RT, Ho GY, Neuhouser ML. Vitamin D intake and lung cancer risk in the Women's Health Initiative. Am J Clin Nutr 2013; 98: 1002-1011.
Higashimoto Y, Ohata M, Nishio K, Iwamoto Y, Fujimoto H, Uetani K, Suruda T, Nakamura Y, Funasako M, Saijo N. 1 alpha, 25-dihydroxyvi- tamin D3 and all-trans-retinoic acid inhibit the growth of a lung cancer cell line. Anticancer Res 1996; 16: 2653-2659.
Nakagawa K, Kawaura A, Kato S, Takeda E, Okano T. 1 alpha, 25-Dihydroxyvitamin D(3) is a preventive factor in the metastasis of lung cancer. Carcinogenesis 2005; 26: 429-440.
Verone-Boyle AR, Shoemaker S, Attwood K, Morrison CD, Makowski AJ, Battaglia S, Hershberger PA. Diet-derived 25-hydroxyvitamin D3 activates vitamin D receptor target gene expression and suppresses EGFR mutant nonsmall cell lung cancer growth in vitro and in vivo. Oncotarget 2015; 7: 995-1013.
Pirianov G, Danielsson C, Carlberg C, James SY, Colston KW. Potentiation by vitamin D analogs of TNFalpha and ceramide-induced apoptosis in MCF-7 cells is associated with activation of cytosolic phospholipase A2. Cell Death Differ 1999; 6: 890-901.
Wang Q, Yang W, Uytingco MS, Christakos S, Wieder R. 1,25-Dihydroxyvitamin D3 and all- trans-retinoic acid sensitize breast cancer cells to chemotherapy-induced cell death. Cancer Res 2000; 60: 2040-2048.
Polar MK, Gennings C, Park M, Gupta MS, Gewirtz DA. Effect of the vitamin D3 analog ILX 23-7553 on apoptosis and sensitivity to fractionated radiation in breast tumor cells and normal human fibroblasts. Cancer Chemother Pharmacol 2003; 51: 415-421.
Mineva ND, Wang X, Yang S, Ying H, Xiao ZX, Holick MF, Sonenshein GE. Inhibition of RelB by 1,25-dihydroxyvitamin D3 promotes sensitivity of breast cancer cells to radiation . J Cell Physiol 2009; 220: 593-599.
Dunlap N, Schwartz GG, Eads D, Cramer SD, Sherk AB, John V, Koumenis C. 1alpha, 25-di- hydroxyvitamin D(3) (calcitriol) and its analogue, 19-nor-1alpha,25(OH)(2)D(2), potentiate the effects of ionising radiation on human prostate cancer cells . Br J Cancer 2003; 89: 746-753.
Sundaram S, Gewirtz DA. The vitamin D3 analog EB 1089 enhances the response of human breast tumor cells to radiation. Radiat Res 1999; 152: 479-486.
Arora S, Tandon C, Tandon S. Evaluation of the cytotoxic effects of CAM therapies: an in vitro study in normal kidney cell lines. Scientific World Journal 2014; 2014: 452892.
Song SJ, Pagel CN, Campbell TM, Pike RN, Mackie EJ. The role of protease-activated receptor-1 in bone healing. Am J Pathol 2005; 166: 857-868.
Tagliabue E, Raimondi S, Gandini S. Vitamin D, Cancer Risk, and Mortality. Adv Food Nutr Res 2015; 75: 1-52.
Ma Y, Trump DL, Johnson CS. Vitamin D in combination cancer treatment. J Cancer 2010; 1: 101-107.
Taneja N, Davis M, Choy JS, Beckett MA, Singh R, Kron SJ, Weichselbaum RR. Histone H2AX phosphorylation as a predictor of radiosensitivity and target for radiotherapy. J Biol Chem 2004; 279: 2273-2280.
Sharma K, Goehe RW, Di X, Hicks MA 2nd, Torti SV, Torti FM, Harada H, Gewirtz DA. A novel cytostatic form of autophagy in sensitization of non-small cell lung cancer cells to radiation by vitamin D and the vitamin D analog, EB 1089. Autophagy 2014; 10: 2346-2361.
Wilson EN, Bristol ML, Di X, Maltese WA, Koterba K, Beckman MJ, Gewirtz DA. A switch between cytoprotective and cytotoxic autophagy in the radiosensitization of breast tumor cells by chloroquine and vitamin D. Horm Cancer 2011; 2: 272-285.
Demasters G, Di X, Newsham I, Shiu R, Gewirtz DA. Potentiation of radiation sensitivity in breast tumor cells by the vitamin D3 analogue, EB 1089, through promotion of autophagy and interference with proliferative recovery. Mol Cancer Ther 2006; 5: 2786-2797.
Roa W, Zhang X, Guo L, Shaw A, Hu X, Xiong Y, Gulavita S, Patel S, Sun X, Chen J, Moore R, Xing JZ. Gold nanoparticle sensitize radiotherapy of prostate cancer cells by regulation of the cell cycle. Nanotechnology 2009; 20: 375101.
Pradier O, Rave-Frank M, Schmidberger H, Bomecke M, Lehmann J, Meden H, Hess CF. Effects of paclitaxel in combination with radiation on human head and neck cancer cells (ZMK-1), cervical squamous cell carcinoma (CaSki), and breast adenocarcinoma cells (MCF-7). J Cancer Res Clin Oncol 1999; 125: 20-27.
Gurrell P. Nursing shortage: psychiatric nursing. Pro Re Nata 1988; 10: 1-4.
Arends MJ, Wyllie AH. Apoptosis: mechanisms and roles in pathology. Int Rev Exp Pathol 1991; 32: 223-254.
Kim HJ, Kim JH, Chie EK, Young PD, Kim IA, Kim IH. DNMOL/LT (DNA methyltransferase) inhibitors radiosensitize human cancer cells by suppressing DNA repair activity. Radiat Oncol 2012; 7: 39.
Qiu H, Yashiro M, Shinto O, Matsuzaki T, Hirakawa K. DNA methyltransferase inhibitor 5-aza-CdR enhances the radiosensitivity of gastric cancer cells. Cancer Sci 2009; 100: 181-188.
Brown JM, Wouters BG. Apoptosis, p53, and tumor cell sensitivity to anticancer agents. Cancer Res 1999; 59: 1391-1399.
Narvaez CJ, Zinser G, Welsh J. Functions of 1al- pha, 25-dihydroxyvitamin D(3) in mammary gland: from normal development to breast cancer. Steroids 2001; 66: 301-308.