Vitamin D levels are strongly associated with genes: overview of twin studies
Genetic Regulation of Vitamin D Levels
CALCIFIED TISSUE INTERNATIONAL, 2012, DOI: 10.1007/s00223-012-9660-z
Published in partnership with the International Osteoporosis Foundation
Zari Dastani, Rui Li and Brent Richards
Vitamin D plays several roles in the body, influencing bone health as well as serum calcium and phosphate levels. Further, vitamin D may modify immune function, cell proliferation, differentiation, and apoptosis. Vitamin D deficiency has been associated with numerous health outcomes, including bone disease, cancer, autoimmune disease, infectious disease, type 1 and type 2 diabetes, hypertension, and heart disease, although it is unclear whether or not these associations are causal. Various twin and family studies have demonstrated moderate to high heritability for circulating vitamin D levels. Accordingly, many studies have investigated the genetic determinants of this hormone. Recent advances in the methodology of large-scale genetic association studies, including coordinated international collaboration, have identified associations of CG, DHCR1, CYP2R1, VDR, and CYP24A1 with serum levels of vitamin D. Here, we review the genetic determinants of vitamin D levels by focusing on new findings arising from candidate gene and genomewide association studies.
The authors have stated that they have no conflict of interest. Z. Dastani • R. Li • B. Richards (&)
Department of Epidemiology, Biostatistics and Occupational Health, Jewish General Hospital, Lady Davis Institute, McGill University, Montreal, QC H3T 1E2, Canada e-mail: brent.richards@mcgill.ca
B. Richards
Departments of Medicine and Human Genetics, Jewish General Hospital, Lady Davis Institute, McGill University, Montreal, QC H3T 1E2, Canada
B. Richards
Department of Twin Research and Genetic Epidemiology, King's College London, London SE1 7EH, UK
Introduction
For decades, vitamin D has been studied intensively due to its association with various diseases of public health importance, which includes bone disease, cancer, autoimmune disease, infectious disease, type 1 and type 2 diabetes, hypertension, and heart disease [1-8]. Many publications have reviewed the relationship between vitamin D and these common diseases [9-15], and this topic is therefore not the subject of this review. Collective evidence from epidemiological, clinical, and experimental studies has demonstrated that both genetic and environmental factors influence vitamin D status. Its heritability has been estimated to be 23-80 % from twin studies [16, 17]. Given this level of heritability, much effort has been dedicated to describing the genetic determinants of this trait. Recent advances in the field of the genetics of vitamin D metabolism have highlighted the importance of several genes. In this review, we first outline the physiology of vitamin D metabolism and examine the evidence of genetic involvement in vitamin D levels, then summarize the approaches that have been used to identify the genes influencing vitamin D levels, and finally comment upon the role of specific variants in candidate genes.
Physiology of Vitamin D
Vitamin D is a fat-soluble prohormone that is crucial for the maintenance of bone and muscle health by promoting the absorption and metabolism of calcium and phosphate.
Fig. 1 Physiology of vitamin D
In addition to food sources such as fatty fish, eggs, fortified milk, and cod liver oil, the human body uses sunlight to synthesize a significant portion of vitamin D requirements. There are two forms of vitamin D: vitamin D2 and D3. The skin synthesizes vitamin D3, or cholecalciferol, after sun exposure, while vitamin D2, or ergocalciferol, is the synthetic form that is often found in fortified food and is derived from plant sources. To become biologically active, the vitamin D originating from dermal production or dietary sources undergoes a series of enzymatic conversions in the liver and kidney (Fig. 1). The hepatic enzyme 25-hydroxylase (CYP2R1) converts vitamin D to 25-hy-droxyvitamin D (25[OH]D), which is the major circulating form of vitamin D [18]. Later, the 25(OH)D-1a-hydroxy-lase (CYP27B1) enzyme, expressed mainly in the kidney and partially in keratinocytes, macrophages, osteoblasts, osteoclasts, dendritic cells, and prostate cells, converts 25(OH)D to 1,25-dihydroxyvitamin D (1a,25[OH]2D) [1923]. 1a,25(OH)2D is the most active form of vitamin D and is responsible for most of its biological actions. These two forms of vitamin D metabolite are commonly measured in serum and have been the target of most genetic studies focusing on vitamin D metabolism. However, 25(OH)D has an almost 1,000-fold greater concentration than 1a,25(OH)2D; also, 25(OH)D has a longer half-life and, hence, is more stable in circulation. Total-body vitamin D stores are best measured by assessing circulating levels of 25(OH)D [24].
Stimulation by parathyroid hormone and inhibition by calcium and phosphate closely regulate the synthesis of 1a,25(OH)2D in the kidney [25-30]. Additionally, calcitonin enhances the production of 1a,25(OH)2D from 25(OH)D in normocalcemic conditions [31, 32]. However, cytokines such as tumor necrosis factor-alpha (TNFa) and interferon-gamma (IFNy) stimulate the production of 1a,25(OH)2D in keratinocytes and macrophages [33]. The vitamin D metabolites are mostly transported by vitamin D binding protein (DBP) (85-88 %) and in part by albumin (12-15 %) [34-36]. Another renal enzyme, 24-a-hydrox-ylase (CYP24), finally hydroxylates both 25(OH)D and 1a,25(OH)2D to initiate degradation of these vitamin D metabolites [37-39] (Fig. 1).
The downstream metabolic activity of the vitamin D pathway is realized through the binding of 1a,25(OH)2Dto the nuclear vitamin D receptor (VDR), which subsequently regulates the expression of genes containing specific DNA sequences known as vitamin D response elements (VDREs) in their promoter regions. 1a,25(OH)2D also has a nongenomic action through binding to membrane receptors to regulate calcium flux in many cells (reviewed elsewhere) [40].
Heritability of Vitamin D
Variability in plasma 25(OH)D is explained by both genetic and environmental factors. The well-replicated, nongenetic determinants include season of measurement, dietary and supplemental vitamin D intake, waist circumference or other measures of obesity such as BMI, and use of hormone replacement therapy (HRT) in women [41-43]. The genetic contribution to variance in vitamin D levels has been investigated in several twin and family studies. Estimates for the heritability of vitamin D levels vary between 23 and 80 %. A study of 1,068 twin pairs (monozygotic [MZ], n = 384; dizygotic [DZ], n = 684) from the TwinsUK estimated heritability to be 43 % for 25(OH)D and 65 % for 1a,25(OH)2D[44]. Orton et al. [45] studied end-of-winter serum 25(OH)D concentrations in 100 adult pairs (MZ, n = 40; DZ, n = 59) from the longitudinal population-based Canadian Collaborative Project on Genetic Susceptibility to multiple sclerosis. They identified the heritability of 25(OH)D levels to be 77 %. Approximately 70 % of the variation in wintertime 25(OH)D concentrations was explained by genetic factors in 510 middle-aged, male twins (310 MZ, 200 DZ) selected from the Vietnam Era Twin Registry [46].
In a cross-sectional study of 1,762 participants of the Framingham Offspring Study, the heritability of plasma 25(OH)D was estimated to be 28.8 % [47]. In a family study of subjects recruited to study asthma, 25(OH)D levels showed a heritability of 80 % and 1a,25(OH)2D, 30 % [16]. Finally, heritability estimates of 25(OH)D and 1a,25(OH)2D were 23 and 16 % in African Americans and Hispanics from the San Luis Valley, Colorado, and 41 and 20 % in Hispanics from San Antonio, Texas, respectively [17]. Table 1 summarizes the heritability estimates. The variation in heritability depends on study design, methods of measuring vitamin D or estimating heritability, and population-specific factors such as allele frequencies and environmental factors. Thus, while there has been considerable variation in the estimation of the heritability of vitamin D levels across these studies, the cumulative evidence suggests that genetic factors play an important role in circulating vitamin D levels.
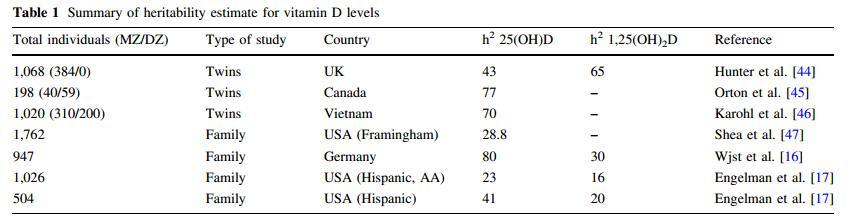
Table 1 Summary of heritability estimate for vitamin D levels
Genetic Studies of Vitamin D
Several studies have reported genetic variants that are associated with alterations in vitamin D levels in humans. These studies are based on different approaches, including linkage studies, the assessment of candidate genes presumed to be involved in vitamin D metabolism, and association analysis from genomewide scans.
Linkage Studies
The classical approach to dissecting the genetics of inherited disease or continuous phenotypes is through linkage analysis, which enables specific genetic intervals across a chromosome to be linked with disease susceptibility. Genetic markers (usually microsatellite markers) are gen-otyped often in families, and linkage analysis determines a locus that segregates with the disease in a pedigree containing affected individuals. The degree of linkage between a trait or disease and genetic markers is measured by the logarithm of odds ratio (LOD) score. The LOD score threshold of 3.3 has been proposed as the threshold of significant linkage [48].
Several approaches of linkage analyses have been developed for complex and quantitative traits. Some investigators use parametric, or model-based, linkage analysis. However, nonparametric, or model-free, linkage approaches have been developed based on identifying a region in affected individuals with shared alleles that are identical by descent (IBD), more than expected by chance. This concept of IBD sharing underpins the identification of the genes for quantitative traits through linkage studies and requires the assumption that if a marker were linked to a gene affecting the trait, two siblings with similar trait values would share IBD more than expected at that locus. This method has been used to identify chromosomal regions that are associated with variability in many diseases and traits, including vitamin D levels.
Although linkage studies have been very effective at identifying genes responsible for Mendelian disorders, they have been mainly unsuccessful in identifying genes involved in quantitative traits, such as vitamin D. A genomewide linkage analysis of family data, including 812 participants recruited for asthma, showed only one region on chromosome 2 at marker D2S2153 (LOD = 3.4) which reached genome-wide significance [16]. Interestingly, this region on chromosome 2 contains genes with known VDREs. Also, this study showed suggestive evidence of linkage for 25(OH)D on chromosome 1 at marker D1S2815 (LOD = 2.9), chromosome 5 at marker D5S2017 (LOD = 2.5), chromosome 6 at marker D6S260 (LOD = 2.1), and chromosome 17 at marker D17S1824 (LOD = 2.5) [16]. A cross-sectional study of 1,762 participants of the Framingham Offspring Study (919 women, mean age 59 years) showed a maximum LOD score for plasma 25(OH)D on chromosome 14 (LOD = 1.16), which did not achieve genomewide significance [47].
The failure of linkage studies can be explained by the polygenic nature of vitamin D regulation, the method of measurement of vitamin D levels, and study design. While linkage has largely failed to identify replicated genetic determinants of vitamin D metabolism, candidate gene approaches and genomewide association studies have identified several reproducible associations with vitamin D levels.
Candidate Gene Studies
Unlike linkage analysis that uses genetic markers coseg-regating with disease status through generations in a family pedigree, candidate gene studies assess whether variation in polymorphic variant frequency associates with variation in a continuous trait, such as vitamin D levels, most often within a group of unrelated individuals. One advantage of this approach is that it allows assignment of this association to a particular DNA base pair variant, rather than a region, such as would be identified through linkage studies.
However, in order to select which regions of the genome to test for association, prior knowledge of the biological mechanism of the trait or disease must be utilized to choose specific genes as the candidates. In a more extensive association study a series of genes involved in a biological pathway related to the targeted trait are studied simultaneously, which is commonly referred to as a pathway analysis. Below we review the loci that have been associated with vitamin D levels through candidate gene studies.
Candidate gene studies have several common limitations: (1) the phenotype characterization is often heterogeneous across studies, (2) sample sizes have tended to be small, and (3) multiple testing correction has not been uniformly applied to such studies, increasing false-positive rates.
It is important to note that the identified associated SNP by candidate gene studies may not be the true disease-causing variant and may reflect the effect of a nearby causal variant, which is in linkage disequilibrium (LD) with the marker. Therefore, replication in separate populations is essential in the identification of candidate genes in order to rule out possible false-positive findings. In view of genomewide association studies, findings with nominally significant p values (<0.05) should be cautiously interpreted.
CYP27B1
CYP27B1, encoding 1 a-hydroxylase which converts 25(OH)D to its active form, 1a,25(OH)2D, has been the cytochrome P-450 gene most strongly associated with vitamin D status. It is located on chromosome 12, at 12q13.1-q13.3, spanning 6.66 kb on the reverse strand. The SNP rs10877012 (C/A) that resides at position 1260 of CYP27B1 was widely explored for the association with 25(OH)D. The rs10877012 C allele was associated with lower levels of 25(OH)D in a study of gestational diabetic patients [49] and in participants from a large cohort study [50]. The effect of the rs10877012 C allele on lower levels of 25(OH)D was also reported in African Americans recently [51]. Although the effect of rs10877012 on 25(OH)D levels has been replicated in candidate gene studies, there is no report on how this single-nucleotide polymorphism (SNP) modulates 25(OH)D levels in serum. However, CYP27B1 functions downstream of circulating 25(OH)D. Therefore, rs10877012, or the causal SNP captured by this SNP, could possibly alter the role of CYP27B1 in metabolic feedback loops or adjust the rate at which 25(OH)D is metabolized [45]. The associations of two other CYP27B1 SNPs, rs4646536 (C/T, +2838) and rs703842 (C/T), with 25(OH)D level were reported in a Canadian multiple sclerosis study [45]. However, associations of these two SNPs were not observed in Hispanics and African Americans [17]. The inconsistency of this association may be due to tight regulation of circulating 1a,25(OH)2D concentrations [52] through 1 a-hydroxyl-ation, the relatively small sample sizes in these studies, or the different ethnicities (Table 2).
CYP2R1
CYP2R1 is responsible for the hydroxylation of vitamin D to 25(OH)D in the first activation step. It is thought to be an important determinant of the vitamin D metabolic pathway as it shows a high affinity for vitamin D [53] and a mis-sense mutation in exon 2 of CYP2R1 leads to vitamin D deficiency [54]. The CYP2R1 gene maps on chromosome 11p15.2, which covers 14.29 kb on the reverse strand. One SNP, rs10741657, which resides in the 5' region of CYP2R1 , was associated with serum 25(OH)D concentrations in a transmission disequilibrium test (TDT) study in a German population [55]. TDT is a family-based association analysis to detect if one of the alleles from a heterozygous parent is overtransmitted to affected offspring compared to the expected ratio of 50:50 [56]. Although this association was not observed in women with gestational diabetes mellitus [49], it was replicated in unrelated individuals from a cohort study [57] and a large study including over 10,000 British individuals [58]. Positive association findings were also reported for two other CYP2R1 SNPs (rs12794714 and rs10766197) in this cohort study [57]. The SNP rs10766197 is located in the 5' flanking region of CYP2R1. The rs10766197 association was observed in a previous study as well [59]. For SNP rs12794714, which causes a synonymous change in the CYP2R1 exon, although no association was found in a previous study involving 133 individuals [55], its relationship with 25(OH)D levels was replicated in a larger study of 2,610 individuals [58] (Table 2).
Table 2 SNPs associated with 25(OH)D
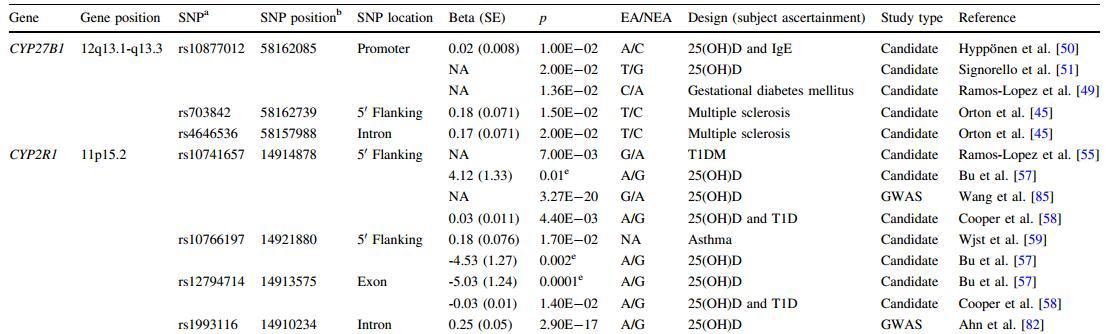
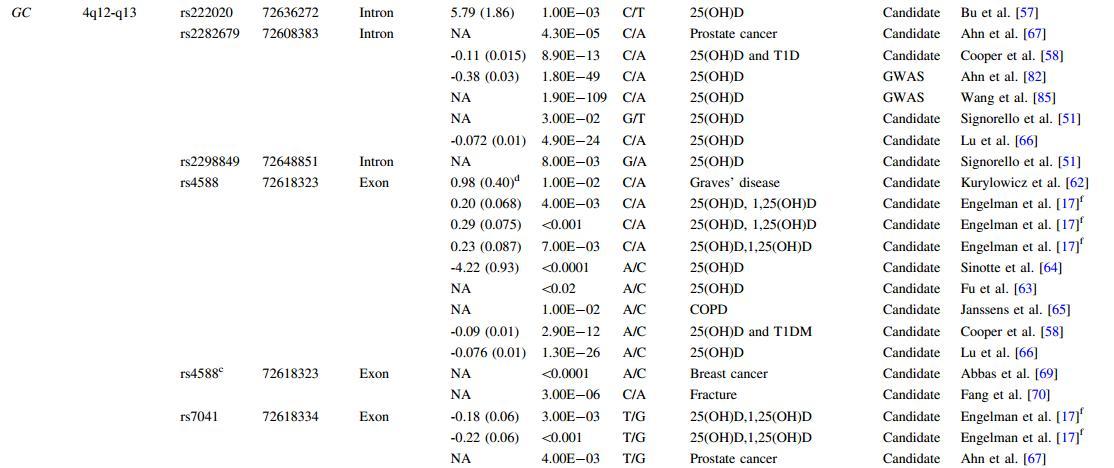

NA = nonavailable, COPD = chronic obstructive pulmonary disease,T1DM = type 1 diabetes mellitus
a -Reported significant SNP
b -Chromosome position in GRCh37.p5 sequence of Genome Build 37.3
c -Haplotype assessment
d -Effect on 25(OH)D deficiency defined as serum concentrations less than or equal to 20 ng/mL
e -Multiple testing corrected p value
f - Associations were reported for three study centers separately in this article
GC
Several studies that investigated the association between vitamin D levels and candidate gene polymorphisms involved in the vitamin D metabolism pathway are reviewed in McGrath et al. [60]. The most widely studied gene is GC, which encodes vitamin D carrier protein, a group-specific component (DBP). GC is located on 4q12-q13 in the human genome, covering 63.84 kb, from 72671237 to 72607403 (NCBI 37, August 2010), on the reverse strand. There are two SNPs in exon 11 of GC, rs7041 (a G/T transversion at codon 416) and rs4588 (a C/A transversion at codon 420). These two SNPs lead to a Glu/Asp amino acid change at codon 416 and a Thr/Lys amino acid chang at codon 420, respectively, and are the most commonly studied variations in DBP [61].
The SNP rs4588 was associated with serum 25(OH)D levels in a Polish population [62]. Carriers of the rs4588 C allele had lower serum levels of 25(OH)D. The association was supported by the lower affinity of DBP variants (the haplotype containing the rs4588 A allele) with higher isoelectric points for 25(OH)D [62]. The effect of rs4588 on 25(OH)D levels has been widely confirmed, with associations described in young Canadian subjects [63], white women [64], Hispanics and African Americans [17], Dutch with chronic obstructive pulmonary disease [65], British [58], and Han Chinese [66]. The T allele at SNP rs7041 was also associated with lower 25(OH)D concentrations consistently through evaluation of the SNP itself [17, 58, 65-68] or the haplotypes derived from both rs4588 and rs7041 [69, 70].
Among other GC SNPs evaluated for an association with 25(OH)D concentrations, several are worthy of note due to their reproducibility across different populations. One is rs2282679, which is situated in intron 12 of this gene and in high LD with rs4588. The association emerged from a study carried out in American males [67] and was replicated in African Americans [51], British [58], and Han Chinese [66], although no association was observed in a small study of Caucasian subjects [57]. An additional variant is rs1155563, another SNP in high LD with rs4588. It was also associated with 25(OH)D level in three independent studies [66-68].
The remaining SNPs listed in Table 2 were associated with 25(OH)D levels in only one study. While positive associations were detected for the SNPs related to either rs4588 or rs7041 (dbSNP IDs of novel SNPs are rs17467825 [68] and rs222035 [68], respectively), the identification of rs16846876 [68] and rs222020 [57], which are not in LD with rs4588 or rs7041, suggests that allelic heterogeneity exists. As well, rs2298849, in the first intron, has been associated with 25(OH)D concentrations in an African American population [51].
In addition to the SNP associations in GC described above, gene-level principal components analysis (PCA) has described an association between this gene and 25(OH)D levels [68]. Thus, both SNP- and gene-level associations have been described between GC and 25(OH)D (Table 2).
VDR
The VDR gene, which spans 63.49 kb on the reverse strand of chromosome 12q12-q14, is another candidate gene. VDR has a large noncoding region containing exons 1F-1C and exons 2-9, which encode a 424-amino acid protein, VDR [71]. The minor allele of VDR SNP rs2228570 (previous dbSNP ID rs10735810), T (f, M4), introduces a FokI (the name of the digestion enzyme used for genotyping) site to exon 2, which leads to a VDR protein that is three amino acids longer by directly introducing a start codon [72]. It influences the activity of the VDR protein [73] and results in a less effective transcriptional activator [74]. The rs2228570 T allele was associated with a higher level of 25(OH)D in a longitudinal population-based study [45]. This association was replicated in another cohort study [72]. In addition, two adjacent SNPs that lie upstream of exon 1A (509 bp distant), rs7139166 (1A-1521) and rs4516035 (1A-1012), were examined for their association with 25(OH)D levels. The rs7139166-rs4516035 (G-A) haplotype showed higher promoter activity, and the rs7139166-rs4516035 (C-G) haplotype was associated with lower circulating levels of 25(OH)D [75]. An intronic SNP, rs10783219, was suggested to be associated with 25(OH)D concentration in a cross-sectional study [17]. These findings await replication (Table 2).
CYP24A1
A third cytochrome P-450 gene to be investigated as a candidate for 25(OH)D concentration regulation is CYP24A1, which encodes the 1a,25(OH)2D inactivation protein. It is located on chromosome 20, at 20q13.2-q13.3, spanning 20.53 kb on the reverse strand. An intronic SNP, rs17219315, was associated with 25(OH)D levels in a family-based study using TDT [59]. This SNP was not included in another association study on CYP24A1 and serum 25(OH)D concentration, in which no significant findings were reported for this gene [67] (Table 2).
Genomewide Association Studies
Rather than focusing on genetic variations in particular genes, over 1 million SNPs across the whole genome can be rapidly genotyped across hundreds or thousands of individuals in genomewide association studies (GWAS), whichtake advantage of the haplotype map of the human genome and advances in array-based genotyping technologies. This approach allows researchers to identify novel associations between disease/phenotype and biological pathways. However, only the common SNPs with the minor allele frequency, generally larger than 1 %, are targeted in GWAS.
The first GWAS of 25(OH)D consisted of 1,012 related individuals from the Framingham Heart Study, which gen-
otyped 70,987 SNPs [76]. No SNPs surpassed the genome–wide significant level of P < 5 x 10-8 in this study. This threshold of genomewide significance is derived from the Bonferroni correction for multiple testing, i.e., 0.05 divided by the number of statistically independent tests. Based on the LD between SNPs, the number of tests is the independent number of SNPs in the European population, which was estimated to be 1 million [77]. In addition, this threshold was previously proposed by a theoretical study [78] and estimated to be robust for a family-wise type I error rate of 0.05 by a permutation test using the Wellcome Trust Case Control Consortium data set [79].
The next GWAS also failed to identify genomewide significant SNPs associated with 25(OH)D concentrations in a study of 229 Hispanic Americans genotyped at 309,200 SNPs [80]. As the effect size of common SNPs is relatively small, GWAS most often requires sample sizes of tens of thousands of individuals to gain enough statistical power to detect reliably disease loci.
Therefore, collaborative efforts to perform comprehensive genomewide meta-analyses have been undertaken to identify such loci. These large-scale international efforts have been made possible through the imputation of un-genotyped SNPs based on the haplotype structure of a reference panel, most often derived from the HapMap or 1000 Genome project. The main objective of this imputation is to expand the density of SNPs assessed across the genome and enable meta-analysis of data from cohorts that have been genotyped on different arrays.
Genomewide significant loci have recently been identified in two large-scale meta-analyses of GWAS. The first study involved 4,501 individuals from five cohorts and 2,221 additional samples in the replication phase, both of European ancestry [81]. Besides confirmation of two candidate genes on the regulation of 25(OH)D levels, GC (rs2282679) and CYP2R1 (rs1993116), a novel association on DHCR7 (rs11234027) was identified (Table 2). DHCR7 encodes a reductase that catalyzes the conversion of 7-dehydrocholesterol to cholesterol in skin [82]. The former is the substrate of the vitamin D synthetic pathway, which is converted to vitamin D induced by UVB radiation found in sunlight. DHCR7 maps to chromosome 11q12-q13, spanning 14.02 kb on the reverse strand. Although it is known that mutations in DHCR7 lead to impaired activity of the gene and consequently accumulation of 7-dehydrocholesterol which results in Smith-Lemli-Opitz syndrome [83], it is the first time that an association has been described between common SNPs in DHCR7 and serum 25(OH)D levels. Further studies are warranted to determine the mechanisms by which genetic variations in this locus regulate 25(OH)D level.
The other GWAS, reported by the SUNLIGHT consortium, involved 33,996 individuals of European descent from 15 cohorts [84]. Given the large sample size of this consortium, the standardized phenotype definitions, and high-quality genotyping, these are likely to be the most reliable results identifying genetic determinants of 25(OH)D levels. Associations on GC (rs2282679), CYP2R1 (rs10741657), and CYP24A1 (rs6013897) were confirmed as well as a novel association identified on DHCR7 (rs12785878) (Table 2). All four susceptibility genes were confirmed in a recent study of 2,610 subjects from a U.K. population [58] (Table 2).
Discussion
The identification of the genetic determinants of any medically relevant trait can, in general, serve three main purposes. The first is to help to explain the physiology of the trait or disease under study, the second is to identify drug targets, and the third is to enable the identification of groups of individuals who may be at risk for disease. Since reduced vitamin D levels have been associated with an increased risk of several diseases of public health importance, the genetic determinants of vitamin D levels may assist in an understanding of the pathophysiology of these diseases or providing drug targets. This, of course, would first require demonstration of a causal link between vitamin D insufficiency, or deficiency, and disease state.
It is important to note that associated SNPs identified by candidate gene studies or GWAS may not be the true disease-causing variants. Only a small subset of SNPs that exist in the genome are genotyped in an association study, which are used as markers of the untyped SNPs. Therefore, the identified associated SNP may reflect the effect of a nearby causal variant, which is in LD with the marker, leading to more efforts such as resequencing of this region to localize the causal SNP. Given that the untyped SNPs in high LD with the marker may span several megabases, dependent on the genetic structure of the region, the findings from a GWAS should be interpreted cautiously.
The main contribution of vitamin D genetics has been to highlight important control points in vitamin D metabolism. While GC and CYP2R1 have been demonstrated to be involved in this pathway and have been the target of relatively intense study, DHCR7 has received little attention as an important determinant of vitamin D levels in humans. The above genetic studies have provided the basis to a better understanding of the role of particularly DHCR7 in vitamin D metabolism.
In addition, it is currently unclear if variation in the GC protein (also called vitamin D binding protein) results in a change in the biologically available amount of vitamin D. This is an important issue to address since common variation in this protein has been associated with a mean change of 25(OH)D levels of up to18 nmol/L, which is larger than the mean change that was associated with vitamin D supplementation [84]. On the other hand, if this large (and common) change in 25(OH)D levels is biologically irrelevant due to a simple change in binding affinity or abundance of GC, then this may have important implications in vitamin D epidemiology.
Another potentially clinically relevant outcome of vitamin D genetics could be an improved understanding of those individuals who would benefit most from supplementation. The change in 25(OH)D levels in response to vitamin D administration is highly variable between individuals [85, 86]. If this change in 25(OH)D levels is determined by genetic factors, it may allow for targeted interventions to those most at risk for vitamin D insufficiency or deficiency.
While twin studies estimate the heritability to be as high as 80 % for vitamin D, the identified common SNPs from GWAS explain up to 4 % of variation in 25(OH)D levels [84]. While this finding is not uncommon in the field of quantitative traits or common disease genetics, it does suggest that most genetic determinants of vitamin D levels have yet to be elucidated. Next-generation DNA sequencing, which will allow for an assessment of rare genetic variants with a minor allele frequency of less than 1 %, may provide an opportunity to identify alleles that have a large effect upon vitamin D levels and may reduce a proportion of this missing heritability.
Norman and Bouillon [87] reviewed the association of vitamin D deficiency with several health outcomes. However, these associations do not imply a clear causal link between vitamin D and disease. Genetic studies may provide a framework to begin to understand whether this relationship is causal through a study design termed Mendelian randomization. Mendelian randomization, which is partially analogous to randomized controlled trials, uses genotype as an instrumental variable to investigate whether the association with the health outcome occurs through its robust association with an intermediate phenotype [88]. Mendelian randomization assumes a random assortment of alleles at the time of conception, therefore eliminating the possible effect of confounders and environmental factors, or reverse causation, on the relation between putative risk factors and disease. Essentially, this method is largely free of confounding since the genotype lies in the causal pathway between the intermediate phenotype (vitamin D levels) and the disease outcome. Application of these methodologies may help to describe the nature of the relationship between circulating vitamin D levels and related common diseases.
In conclusion, in addition to the key environmental determinants of vitamin D levels, genetic factors play an important role in modifying the abundance of this clinically relevant hormone. Candidate gene studies, in particular GWAS, have been successful at identifying proteins in humans that are important determinants of 25(OH)D concentrations. Whole-genome sequencing and Mendelian randomization may provide new insights into vitamin D metabolism and may help to delineate its relationship with common disease.
References
Bouillon R, Bischoff-Ferrari H, Willett W (2008) Vitamin D and health: perspectives from mice and man. J Bone Miner Res 23:974-979
Hypponen E, Laara E, Reunanen A, Jarvelin MR, Virtanen SM (2001) Intake of vitamin D and risk of type 1 diabetes: a birth-cohort study. Lancet 358:1500-1503
Zipitis CS, Akobeng AK (2008) Vitamin D supplementation in early childhood and risk of type 1 diabetes: a systematic review and meta-analysis. Arch Dis Child 93:512-517
Wang TJ, Pencina MJ, Booth SL, Jacques PF, Ingelsson E, Lanier K, Benjamin EJ, D'Agostino RB, Wolf M, Vasan RS (2008) Vitamin D deficiency and risk of cardiovascular disease. Circulation 117:503-511
Giovannucci E, Liu Y, Rimm EB, Hollis BW, Fuchs CS, Stampfer MJ, Willett WC (2006) Prospective study of predictors of vitamin D status and cancer incidence and mortality in men. J Natl Cancer Inst 98:451-459
Martinez ME, Giovannucci EL, Colditz GA, Stampfer MJ, Hunter DJ, Speizer FE, Wing A, Willett WC (1996) Calcium, vitamin D, and the occurrence of colorectal cancer among women. J Natl Cancer Inst 88:1375-1382
Garland CF, Garland FC, Gorham ED, Lipkin M, Newmark H, Mohr SB, Holick MF (2006) The role of vitamin D in cancer prevention. Am J Public Health 96:252-261
John EM, Schwartz GG, Dreon DM, Koo J (1999) Vitamin D and breast cancer risk: the NHANES I Epidemiologic follow-up study, 1971-1975 to 1992. National health and nutrition examination survey. Cancer Epidemiol Biomarkers Prev 8:399-406
Temmerman JC (2011) Vitamin D and cardiovascular disease. J Am Coll Nutr 30:167-170
Melamed ML, Manson JE (2011) Vitamin D and cardiovascular disease and cancer: not too much and not too little? The need for clinical trials. Womens Health (Lond Engl) 7:419-424
Van Belle TL, Gysemans C, Mathieu C (2011) Vitamin D in autoimmune, infectious and allergic diseases: a vital player? Best Pract Res Clin Endocrinol Metab 25:617-632
Lips P, van Schoor NM (2011) The effect of vitamin D on bone and osteoporosis. Best Pract Res Clin Endocrinol Metab 25:585-591
Vanoirbeek E, Krishnan A, Eelen G, Verlinden L, Bouillon R, Feldman D, Verstuyf A (2011) The anti-cancer and anti-inflammatory actions of 1,25(OH)D. Best Pract Res Clin Endocrinol Metab 25:593-604
Mitri J, Muraru MD, Pittas AG (2011) Vitamin D and type 2 diabetes: a systematic review. Eur J Clin Nutr 65:1005-1015
Elamin MB, Abu Elnour NO, Elamin KB, Fatourechi MM, Al-katib AA, Almandoz JP, Liu H, Lane MA, Mullan RJ, Hazem A, Erwin PJ, Hensrud DD, Murad MH, Montori VM (2011) Vitamin D and cardiovascular outcomes: a systematic review and meta-analysis. J Clin Endocrinol Metab 96:1931-1942
Wjst M, Altmuller J, Braig C, Bahnweg M, Andre E (2007) A genome-wide linkage scan for 25-OH-D3 and 1,25-(OH)2D3 serum levels in asthma families. J Steroid Biochem Mol Biol 103:799-802
Engelman CD, Fingerlin TE, Langefeld CD, Hicks PJ, Rich SS, Wagenknecht LE, Bowden DW, Norris JM (2008) Genetic and Z. Dastani et al.: Genetic Regulation of Vitamin D environmental determinants of 25-hydroxyvitamin D and 1,25-dihydroxyvitamin D levels in Hispanic and African Americans. J Clin Endocrinol Metab 93:3381-3388
Haddad JG, Matsuoka LY, Hollis BW, Hu YZ, Wortsman J 1993) Human plasma transport of vitamin D after its endogenous synthesis. J Clin Invest 91:2552-2555
Zehnder D, Bland R, Williams MC, McNinch RW, Howie AJ, Stewart PM, Hewison M (2001) Extrarenal expression of 25-hydroxyvitamin d3-1 alpha-hydroxylase. J Clin Endocrinol
Metab 86:888-894
Flanagan JN, Wang L, Tangpricha V, Reichrath J, Chen TC, Holick MF (2003) Regulation of the 25-hydroxyvitamin D-1alpha-hydroxylase gene and its splice variant. Recent Results Cancer Res 164:157-167
Wang L, Whitlatch LW, Flanagan JN, Holick MF, Chen TC (2003) Vitamin D autocrine system and prostate cancer. Recent Results Cancer Res 164:223-237
Fritsche J, Mondal K, Ehrnsperger A, Andreesen R, Kreutz M (2003) Regulation of 25-hydroxyvitamin D3-1alpha-hydroxylase and production of 1alpha,25-dihydroxyvitamin D3 by human dendritic cells. Blood 102:3314-3316
Kogawa M, Findlay DM, Anderson PH, Ormsby R, Vincent C, Morris HA, Atkins GJ (2010) Osteoclastic metabolism of 25(OH)-vitamin D3: a potential mechanism for optimization of bone resorption. Endocrinology 151:4613-4625
Lips P (2007) Relative value of 25(OH)D and 1,25(OH)2D measurements. J Bone Miner Res 22:1668-1671
Rasmussen H, Wong M, Bikle D, Goodman DB (1972) Hormonal control of the renal conversion of 25-hydroxycholecalciferol to 1,25-dihydroxycholecalciferol. J Clin Invest 51:2502-2504
Bikle DD, Murphy EW, Rasmussen H (1975) The ionic control of 1,25-dihydroxyvitamin D3 synthesis in isolated chick renal mitochondria. The role of calcium as influenced by inorganic phosphate and hydrogen-ion. J Clin Invest 55:299-304
Rost CR, Bikle DD, Kaplan RA (1981) In vitro stimulation of 25-hydroxycholecalciferol 1alpha-hydroxylation by parathyroid hormone in chick kidney slices: evidence for a role for adenosine 3',5'-monophosphate. Endocrinology 108:1002-1006
Hughes MR, Brumbaugh PF, Hussler MR, Wergedal JE, Baylink
DJ (1975) Regulation of serum 1alpha,25-dihydroxyvitamin D3 by calcium and phosphate in the rat. Science 190:578-580
Portale AA, Halloran BP, Morris RC Jr (1987) Dietary intake of phosphorus modulates the circadian rhythm in serum concentration of phosphorus. Implications for the renal production of 1,25-dihydroxyvitamin D. J Clin Invest 80:1147-1154
Condamine L, Menaa C, Vrtovsnik F, Friedlander G, Garabedian M (1994) Local action of phosphate depletion and insulin-like growth factor 1 on in vitro production of 1,25-dihydroxyvitamin D by cultured mammalian kidney cells. J Clin Invest 94:1673-1679
Jaeger P, Jones W, Clemens TL, Hayslett JP (1986) Evidence that calcitonin stimulates 1,25-dihydroxyvitamin D production and intestinal absorption of calcium in vivo. J Clin Invest 78:456-461
Shinki T, Ueno Y, DeLuca HF, Suda T (1999) Calcitonin is a major regulator for the expression of renal 25-hydroxyvitamin D3-1alpha-hydroxylase gene in normocalcemic rats. Proc Natl Acad Sci USA 96:8253-8258
Bikle DD, Pillai S (1993) Vitamin D, calcium, and epidermal differentiation. Endocr Rev 14:3-19
Cooke NE, Haddad JG (1989) Vitamin D binding protein (Gc-globulin). Endocr Rev 10:294-307
Bikle DD, Gee E, Halloran B, Haddad JG (1984) Free 1,25-dihydroxyvitamin D levels in serum from normal subjects, pregnant subjects, and subjects with liver disease. J Clin Invest 74:1966-1971
Bikle DD, Siiteri PK, Ryzen E, Haddad JG (1985) Serum protein binding of 1,25-dihydroxyvitamin D: a reevaluation by direct measurement of free metabolite levels. J Clin Endocrinol Metab 61:969-975
Kawashima H, Torikai S, Kurokawa K (1981) Localization of 25-hydroxyvitamin D3 1alpha-hydroxylase and 24-hydroxylase along the rat nephron. Proc Natl Acad Sci USA 78:1199-1203
Zierold C, Darwish HM, DeLuca HF (1994) Identification of a vitamin D-response element in the rat calcidiol (25-hydroxyvitamin D3) 24-hydroxylase gene. Proc Natl Acad Sci USA 91:900-902
Zehnder D, Bland R, Walker EA, Bradwell AR, Howie AJ, Hewison M, Stewart PM (1999) Expression of 25-hydroxyvita-min D3-1alpha-hydroxylase in the human kidney. J Am Soc Nephrol 10:2465-2473
Lips P (2006) Vitamin D physiology. Prog Biophys Mol Biol 92:4-8
Burnand B, Sloutskis D, Gianoli F, Cornuz J, Rickenbach M, Paccaud F, Burckhardt P (1992) Serum 25-hydroxyvitamin D: distribution and determinants in the Swiss population. Am J Clin Nutr 56:537-542
Jacques PF, Felson DT, Tucker KL, Mahnken B, Wilson PW, Rosenberg IH, Rush D (1997) Plasma 25-hydroxyvitamin D and its determinants in an elderly population sample. Am J Clin Nutr 66:929-936
Rossouw JE, Anderson GL, Prentice RL, LaCroix AZ, Kooper-berg C, Stefanick ML, Jackson RD, Beresford SA, Howard BV, Johnson KC, Kotchen JM, Ockene J (2002) Risks and benefits of estrogen plus progestin in healthy postmenopausal women: principal results from the Women's health initiative randomized controlled trial. JAMA 288:321-333
Hunter D, De Lange M, Snieder H, MacGregor AJ, Swaminathan R, Thakker RV, Spector TD (2001) Genetic contribution to bone metabolism, calcium excretion, and vitamin D and parathyroid hormone regulation. J Bone Miner Res 16:371-378
Orton SM, Morris AP, Herrera BM, Ramagopalan SV, Lincoln MR, Chao MJ, Vieth R, Sadovnick AD, Ebers GC (2008) Evidence for genetic regulation of vitamin D status in twins with multiple sclerosis. Am J Clin Nutr 88:441-447
Karohl C, Su S, Kumari M, Tangpricha V, Veledar E, Vaccarino V, Raggi P (2010) Heritability and seasonal variability of vitamin D concentrations in male twins. Am J Clin Nutr 92:1393-1398
Shea MK, Benjamin EJ, Dupuis J, Massaro JM, Jacques PF, D'Agostino RB Sr, Ordovas JM, O'Donnell CJ, Dawson-Hughes B, Vasan RS, Booth SL (2009) Genetic and non-genetic correlates of vitamins K and D. Eur J Clin Nutr 63:458-464
Lander E, Kruglyak L (1995) Genetic dissection of complex traits: guidelines for interpreting and reporting linkage results. Nat Genet 11:241-247
Ramos-Lopez E, Kahles H, Weber S, Kukic A, Penna-Martinez M, Badenhoop K, Louwen F (2008) Gestational diabetes mellitus and vitamin D deficiency: genetic contribution of CYP27B1 and CYP2R1 polymorphisms. Diabetes Obes Metab 10:683-685
Hypponen E, Berry DJ, Wjst M, Power C (2009) Serum 25-hy-droxyvitamin D and IgE—a significant but nonlinear relationship. Allergy 64:613-620
Signorello LB, Shi J, Cai Q, Zheng W, Williams SM, Long J, Cohen SS, Li G, Hollis BW, Smith JR, Blot WJ (2011) Common variation in vitamin D pathway genes predicts circulating 25-hydroxyvitamin D levels among African Americans. PLoS
ONE 6:e28623
Berry D, Hypponen E (2011) Determinants of vitamin D status: focus on genetic variations. Curr Opin Nephrol Hypertens 20: 331-336
Shinkyo R, Sakaki T, Kamakura M, Ohta M, Inouye K (2004) Metabolism of vitamin D by human microsomal CYP2R1. Bio-chem Biophys Res Commun 324:451-457
Cheng JB, Levine MA, Bell NH, Mangelsdorf DJ, Russell DW (2004) Genetic evidence that the human CYP2R1 enzyme is a 1 S Z. Dastani et al.: Genetic Regulation of Vitamin D key vitamin D 25-hydroxylase. Proc Natl Acad Sci USA 101:
7711-7715
Ramos-Lopez E, Bruck P, Jansen T, Herwig J, Badenhoop K (2007) CYP2R1 (vitamin D 25-hydroxylase) gene is associated with susceptibility to type 1 diabetes and vitamin D levels in Germans. Diabetes Metab Res Rev 23:631-636
Spielman RS, McGinnis RE, Ewens WJ (1993) Transmission test for linkage disequilibrium: the insulin gene region and insulin-dependent diabetes mellitus (IDDM). Am J Hum Genet 52: 506-516
Bu FX, Armas L, Lappe J, Zhou Y, Gao G, Wang HW, Recker R, Zhao LJ (2010) Comprehensive association analysis of nine candidate genes with serum 25-hydroxyvitamin D levels among healthy Caucasian subjects. Hum Genet 128:549-556
Cooper JD, Smyth DJ, Walker NM, Stevens H, Burren OS, Wallace C, Greissl C, Ramos-Lopez E, Hypponen E, Dunger DB, Spector TD, Ouwehand WH, Wang TJ, Badenhoop K, Todd JA
(2011) Inherited variation in vitamin D genes is associated with predisposition to autoimmune disease type 1 diabetes. Diabetes 60:1624-1631
Wjst M, Altmuller J, Faus-Kessler T, Braig C, Bahnweg M, Andre E (2006) Asthma families show transmission disequilibrium of gene variants in the vitamin D metabolism and signalling pathway. Respir Res 7:60
McGrath JJ, Saha S, Burne TH, Eyles DW (2010) A systematic
review of the association between common single nucleotide polymorphisms and 25-hydroxyvitamin D concentrations. J Steroid Biochem Mol Biol 121:471-477
Braun A, BichlmaierR, Cleve H (1992) Molecularanalysis ofthe gene for the human vitamin-D-binding protein (group-specific component): allelic differences of the common genetic GC types. Hum Genet 89:401-406
Kurylowicz A, Ramos-Lopez E, Bednarczuk T, Badenhoop K (2006) Vitamin D-binding protein (DBP) gene polymorphism is associated with Graves' disease and the vitamin D status in a Polish population study. Exp Clin Endocrinol Diabetes 114: 329-335
Fu L, Wagner D, Hanwell HEC, Sidhom G, Zhu J, Wong BYL, Vieth R, Cole DEC (2008) Functional T436 K SNP in DBP is a predictor of serum 25(OH)D: a replicate study in a young healthy adult population. Clin Biochem 41:1267
Sinotte M, Diorio C, Berube S, Pollak M, Brisson J (2009) Genetic polymorphisms of the vitamin D binding protein and plasma concentrations of 25-hydroxyvitamin D in premenopausal
women. Am J Clin Nutr 89:634-640
Janssens W, Bouillon R, Claes B, Carremans C, Lehouck A, Buysschaert I, Coolen J, Mathieu C, Decramer M, Lambrechts D (2010) Vitamin D deficiency is highly prevalent in COPD and correlates with variants in the vitamin D-binding gene. Thorax 65:215-220
Lu L, Sheng H, Li H, Gan W, Liu C, Zhu J, Loos RJ, Lin X (2012) Associations between common variants in GC and DHCR7/NADSYN1 and vitamin D concentration in Chinese Hans. Hum Genet 131:505-512
Ahn J, Albanes D, Berndt SI, Peters U, Chatterjee N, Freedman ND, Abnet CC, Huang WY, Kibel AS, Crawford ED, Weinstein SJ, Chanock SJ, Schatzkin A, Hayes RB (2009) Vitamin D-
related genes, serum vitamin D concentrations and prostate cancer risk. Carcinogenesis 30:769-776
- Hibler EA, Hu C, Jurutka PW, Martinez ME, Jacobs ET (2012)
Polymorphic variation in the GC and CASR genes and associations with vitamin D metabolite concentration and metachronous colorectal neoplasia. Cancer Epidemiol Biomarkers Prev 21:
368-375
- Abbas S, Linseisen J, Slanger T, Kropp S, Mutschelknauss EJ,
Flesch-Janys D, Chang-Claude J (2008) The Gc2 allele of the vitamin D binding protein is associated with a decreased post-menopausal breast cancer risk, independent of the vitamin D status. Cancer Epidemiol Biomarkers Prev 17:1339-1343
Fang Y, van Meurs JB, Arp P, van Leeuwen JP, Hofman A, Pols HA, Uitterlinden AG (2009) Vitamin D binding protein genotype and osteoporosis. Calcif Tissue Int 85:85-93
Uitterlinden AG, Fang Y, Van Meurs JB, Pols HA, Van Leeuwen JP (2004) Genetics and biology of vitamin D receptor polymorphisms. Gene 338:143-156
Smolders J, Damoiseaux J, Menheere P, Tervaert JW, Hupperts R (2009) Fok-I vitamin D receptor gene polymorphism (rs10735810) and vitamin D metabolism in multiple sclerosis.
J Neuroimmunol 207:117-121
Haussler MR, Whitfield GK, Haussler CA, Hsieh JC, Thompson PD, Selznick SH, Dominguez CE, Jurutka PW (1998) The nuclear vitamin D receptor: biological and molecular regulatory properties revealed. J Bone Miner Res 13:325-349
Arai H, Miyamoto KI, Yoshida M, Yamamoto H, Taketani Y, Morita K, Kubota M, Yoshida S, Ikeda M, Watabe F, Kanemasa Y, Takeda E (2001) The polymorphism in the caudal-related homeodomain protein Cdx-2 binding element in the human vitamin D receptor gene. J Bone Miner Res 16:1256-1264
d'Alesio A, Garabedian M, Sabatier JP, Guaydier-Souquieres G, Marcelli C, Lemacon A, Walrant-Debray O, Jehan F (2005) Two single-nucleotide polymorphisms in the human vitamin D receptor promoter change protein-DNA complex formation and are associated with height and vitamin D status in adolescent
girls. Hum Mol Genet 14:3539-3548
- Benjamin EJ, Dupuis J, Larson MG, Lunetta KL, Booth SL, Govindaraju DR, Kathiresan S, Keaney JF Jr, Keyes MJ, Lin JP, Meigs JB, Robins SJ, Rong J, Schnabel R, Vita JA, Wang TJ, Wilson PW, Wolf PA, Vasan RS (2007) Genome-wide association with select biomarker traits in the Framingham Heart Study.
BMC Med Genet 8(Suppl 1):S11
- Frazer KA, Ballinger DG, Cox DR, Hinds DA, Stuve LL, Gibbs RA, Belmont JW, Boudreau A, Hardenbol P, Leal SM, Pasternak S, Wheeler DA, Willis TD, Yu F, Yang H, Zeng C, Gao Y, Hu H, Hu W, Li C, Lin W, Liu S, Pan H, Tang X, Wang J, Wang W, Yu J, Zhang B, Zhang Q, Zhao H, Zhou J, Gabriel SB, Barry R,
Blumenstiel B, Camargo A, Defelice M, Faggart M, Goyette M, Gupta S, Moore J, Nguyen H, Onofrio RC, Parkin M, Roy J, Stahl E, Winchester E, Ziaugra L, Altshuler D, Shen Y, Yao Z, Huang W, Chu X, He Y, Jin L, Liu Y, Sun W, Wang H, Wang Y, Xiong X, Xu L, Waye MM, Tsui SK, Xue H, Wong JT, Galver LM, Fan JB, Gunderson K, Murray SS, Oliphant AR, Chee MS, Montpetit A, Chagnon F, Ferretti V, Leboeuf M, Olivier JF, Phillips MS, Roumy S, Sallee C, Verner A, Hudson TJ, Kwok PY, Cai D, Koboldt DC, Miller RD, Pawlikowska L, Taillon-Miller P, Xiao M, Tsui LC, Mak W, Song YQ, Tam PK, Nakamura Y, Kaw-aguchi T, Kitamoto T, Morizono T, Nagashima A, Ohnishi Y, Sekine A, Tanaka T, Tsunoda T et al (2007) A second generation human haplotype map of over 3.1 million SNPs. Nature 449: 851-861
Risch N, Merikangas K (1996) The future of genetic studies of complex human diseases. Science 273:1516-1517
Dudbridge F, Gusnanto A (2008) Estimation of significance thresholds for genomewide association scans. Genet Epidemiol 32:227-234
Engelman CD, Meyers KJ, Ziegler JT, Taylor KD, Palmer ND, Haffner SM, Fingerlin TE, Wagenknecht LE, Rotter JI, Bowden DW, Langefeld CD, Norris JM (2010) Genome-wide association study of vitamin D concentrations in Hispanic Americans: the IRAS family study. J Steroid Biochem Mol Biol 122:186-192
Ahn J, Yu K, Stolzenberg-Solomon R, Simon KC, McCullough ML, Gallicchio L, Jacobs EJ, Ascherio A, Helzlsouer K, Jacobs KB, Li Q, Weinstein SJ, Purdue M, Virtamo J, Horst R, Wheeler Z. Dastani et al.: Genetic Regulation of Vitamin D W, Chanock S, Hunter DJ, Hayes RB, Kraft P, Albanes D (2010) Genome-wide association study of circulating vitamin D levels. Hum Mol Genet 19:2739-2745
Waterham HR, Wanders RJ (2000) Biochemical and genetic aspects of 7-dehydrocholesterol reductase and Smith-Lemli-Opitz syndrome. Biochim Biophys Acta 1529:340-356
Tint GS, Irons M, Elias ER, Batta AK, Frieden R, Chen TS, Salen G (1994) Defective cholesterol biosynthesis associated with the Smith-Lemli-Opitz syndrome. N Engl J Med 330:107-113
Wang TJ, Zhang F, Richards JB, Kestenbaum B, van Meurs JB, Berry D, Kiel DP, Streeten EA, Ohlsson C, Koller DL, Peltonen L, Cooper JD, O'Reilly PF, Houston DK, Glazer NL, Vandenput L, Peacock M, Shi J, Rivadeneira F, McCarthy MI, Anneli P, de Boer IH, Mangino M, Kato B, Smyth DJ, Booth SL, Jacques PF, Burke GL, Goodarzi M, Cheung CL, Wolf M, Rice K, Goltzman D, Hidiroglou N, Ladouceur M, Wareham NJ, Hocking LJ, Hart D, Arden NK, Cooper C, Malik S, Fraser WD, Hartikainen AL, Zhai G, Macdonald HM, Forouhi NG, Loos RJ, Reid DM, Hakim A, Dennison E, Liu Y, Power C, Stevens HE, Jaana L, Vasan RS, Soranzo N, Bojunga J, Psaty BM, Lorentzon M, Foroud T, Harris TB, Hofman A, Jansson JO, Cauley JA, Uitterlinden AG, Gibson Q, Jarvelin MR, Karasik D, Siscovick DS, Econs MJ, Kritchev-sky SB, Florez JC, Todd JA, Dupuis J, Hypponen E, Spector TD (2010) Common genetic determinants of vitamin D insufficiency: a genome-wide association study. Lancet 376:180-188
Meyer HE, Smedshaug GB, Kvaavik E, Falch JA, Tverdal A, Pedersen JI (2002) Can vitamin D supplementation reduce the risk of fracture in the elderly? A randomized controlled trial. J Bone Miner Res 17:709-715
Dawson-Hughes B, Dallal GE, Krall EA, Sadowski L, Sahyoun N, Tannenbaum S (1990) A controlled trial of the effect of calcium supplementation on bone density in postmenopausal women. N Engl J Med 323:878-883
Norman AW, Bouillon R (2010) Vitamin D nutritional policy needs a vision for the future. Exp Biol Med (Maywood) 235: 1034-1045
Lawlor DA, Harbord RM, Sterne JA, Timpson N, Davey Smith G (2008) Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med 27: 1133-1163 1 S
PDF is attached at the bottom of this page
See also VitaminDWiki
Twins reduction in vitamin D in the winter or summer - Nov 2010
40 percent less likely to get adequate vitamin D response if have certain genes – Jan 2013
Genes account for less than 18 percent of variation in vitamin D levels – Jan 2013
229 Genes related to vitamin D - Aug 2010 which contains the following graph
See also web
- Clinical trials of Genes and Vitamin D 70 as of May 2014