Vaccines are plan A for COVID-19 Immunity (no plan B )
COVID-19 and the Path to Immunity
JAMA. 2020;324(13):1279-1281. doi:10.1001/jama.2020.16656
David S. Stephens, MD1; M. Juliana McElrath, MD, PhD2
Corresponding Author: David S. Stephens, MD, Emory University School of Medicine, Robert W. Woodruff Health Sciences Center, Emory University, 1440 Clifton Rd NE, #420, Atlanta, GA 30322 (dstep01@emory.edu).
Vitamin D should be plan B
COVID-19 treated by Vitamin D - studies, reports, videos
{include}
15 studies indicating that Vitamin D fights COVID-19 - Dr. Grimes Oct 9, 2020
Vitamin D could knock out COVID-19 in 3 months – Dr. Matthews interview Oct 2020
100,000 dollar bet that Vitamin D is effective in reducing the severity of Covid-19 - Sept 23, 2020
The Covid-19 pandemic is a Vitamin D Deficiency problem and is easy to solve – June 2020
Perhaps the Vitamin D deficiency pandemic caused the COVID-19 pandemic - Aug 17, 2020
Prudent to consider that Vitamin D has a role in COVID-19 – meta-analysis – Aug 7, 2020
Vitamin D fought viruses before COVID-19
How Vitamin D helps fight the cold, flu and other viruses - June 2019
Vitamin D 10 x better than Flu Vaccine if you have very low vitamin D – Feb 2017
Influenza virus might be prevented and treated by Vitamin D, if no vaccination – Aug 2018
7X less risk of influenza if Vitamin D levels higher than 30 ng – Oct 2017
-
Vitamin D proven to fight more than virus
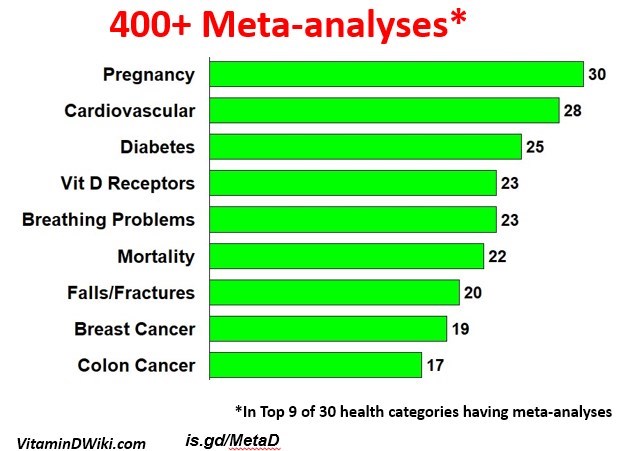
92 Health Problems were proven to be fought by Vitamin D as of Sept 2020
Much more likely to live longer if higher vitamin D – 27,000 seniors Feb 2017 13 meta-analysis
Top 10 causes of death - low vitamin D is associated with every cause - Nov 2018
Many meta-analysis have agreed that Vitamin D helps (as of April 2019)
📄 Download the PDF from VitaminDWiki
The emergence of adaptive immunity in response to the novel Betacoronavirus, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), occurs within the first 7 to 10 days of infection.1-3 Understanding the key features and evolution of B-cell– and T-cell–mediated adaptive immunity to SARS-CoV-2 is essential in forecasting coronavirus disease 2019 (COVID-19) outcomes and for developing effective strategies to control the pandemic. Ascertaining long-term B-cell and T-cell immunological memory against SARS-CoV-2 is also critical to understanding durable protection.
A robust memory B-cell and plasmablast expansion is detected early in infection,2,4 with secretion of serum IgM and IgA antibodies by day 5 to 7 and IgG by day 7 to 10 from the onset of symptoms. In general, serum IgM and IgA titers decline after approximately 28 days (Figure), and IgG titers peak at approximately 49 days. Simultaneously, SARS-CoV-2 activates T cells in the first week of infection, and virus-specific memory CD4+ cells and CD8+ T cells reportedly peak within 2 weeks but remain detectable at lower levels for 100 or more days of observation. Grifoni et al1 and others5,6 have identified SARS-CoV-2–specific memory CD4+ T cells in up to 100% and CD8+ T cells in approximately 70% of patients recovering from COVID-19. Although severe COVID-19 is characterized by high-viral titers, dysregulated innate inflammatory cytokine and chemokine responses and prolonged lymphopenia, antibody-dependent enhancement or dominant CD4+ TH2-type cytokines (eg, IL-4, IL-5, IL-13) do not appear to contribute to acute COVID-19 severity.
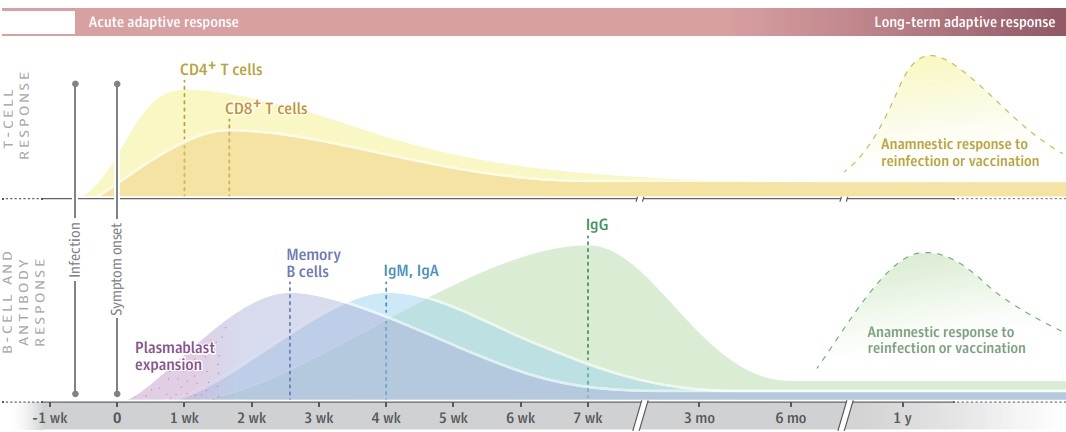
Figure. Adaptive Immunity to Coronavirus Disease 2019
Generalized model of T-cell and B-cell (plasmablast, antibody) responses to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection projected over 1 year following infection. Neutralizing antibodies, memory B cells, and CD4+ and CD8+ memory T cells to SARS-CoV-2, which are generated by infection, vaccination, or after reexposure, are key to the path to immunity. The dotted lines represent peak B-cell, T-cell, and antibody responses following infection.
The magnitude of the antibody and T-cell responses can differ and be discordant among individuals and is influenced by disease severity (asymptomatic, mild, moderate, or severe). The immune correlates of protection are not yet defined for COVID-19, but neutralizing antibodies, especially those that recognize the viral receptor binding domain (RBD) and other epitopes on the spike protein that prevent subsequent angiotensin-converting enzyme II receptor binding, membrane fusion, and viral entry, is one path to immunity. The magnitude of the anti–SARS-CoV-2 IgG and IgA titers to the spike protein correlates in convalescing patients with CD4+ T-cell responses1; and the magnitude of IgG1 and IgG3 RBD enzyme-linked immunosorbent assay (ELISA) titers correlates strongly with viral neutralization.2,3
The generation of neutralizing antibodies directed at the spike protein is a basis of multiple human vaccines in clinical trials7 to counteract SARS-CoV-2, and virus neutralization is the basis of potential therapeutic and preventive monoclonal antibodies also currently in human clinical trials. Such virus neutralizing antibodies are protective in animal models of SARS-CoV-2 infection. Potent neutralizing antibodies and TH1-biased CD4+ T-cell responses to the spike protein protect against SARS-CoV-2 infection in the lungs and nasal mucosa of nonhuman primates without evidence of immunopathological changes.8 The RBD region of the SARS-CoV-2 spike protein shows little sequence homology to the seasonal coronaviruses.2 Although variants in the SARS-CoV-2 spike protein (eg, D614G) may be a concern, SARS-CoV-2 has a low variation rate and so far, such variants have not been shown to reduce the recognition of RBD epitopes important for antibody neutralization.
Recent reports have demonstrated a decline in IgG neutralizing antibodies to SARS-CoV-2 in convalescence, raising apprehension of susceptibility to reinfection. Antibody levels always decline after the acute phase of infection because most of the plasmablasts, the “effector” response of B cells, induced during the first weeks after infection are short-lived. A similar pattern is seen with the effector CD8+ T-cell response. After this reduction, serological memory is then maintained by the smaller number of long-lived plasma cells that reside in the bone marrow and constitutively secrete antibody in the absence of antigen. The antibody recall response comes from this pool of memory B cells that are also long-lived. In fact, rare circulating memory cells have been shown to produce highly potent neutralizing antibodies when serum neutralizing titers are low.3 Thus, an early decline of neutralizing antibody levels should not be of concern. The key is at what levels the antibody titers stabilize after natural infection or vaccination. This represents the generation of long-lived plasma cells to protect against subsequent infection.
About one-third of patients who have recovered from COVID-19 do have antibodies with low anti-RBD titers and low viral neutralizing activity, especially among those who have had mild or asymptomatic disease. Given the wide range of clinical disease, this variability in the antibody responses among patients with COVID-19 is expected. Antigen burden is a major driver of the magnitude of the response, as notably the highest neutralizing antibody titers are found in severe disease,3 but other factors also could be involved. Based on models of immunity and reinfection dynamics of the common cold coronaviruses, human coronavirus (HCoV) OC43, HCoV 229E, and HCoV HKU1, limited protective immunity to SARS-CoV-2 has been suggested. In a study involving a human challenge to a circulating coronavirus (HCoV 229E), IgG and IgA antibodies waned over the first year after viral nasal challenge suggesting that protection against repeated infections with common cold coronaviruses lasts only 1 or 2 years.9 However, following experimental rechallenge with the same HCoV 229E strain at 1 year, no individuals who had been previously infected developed a cold and all had a shorter duration of detectable virus shedding. Thus, at least strain-specific immunity to clinical coronavirus disease may be preserved despite rapid waning of antibodies. In nonhuman primates, SARS-CoV-2 infection protects against reinfection.10 More than 8 months into the outbreak and after millions of infections globally, anecdotal case reports of reinfection mostly after initial mild COVID-19 illness are appearing. Although the complete immune profile of these individuals is not clear, reinfection with SARS-CoV-2 suggests that the natural human immune response may not provide sterilizing immunity but that it may shorten viral shedding, reduce spread, and prevent disease.
SARS-CoV-2–specific CD4+ and CD8+ memory T cells are also generated in asymptomatic to severe disease,1,5,6 which exhibit cytotoxic activities and express antiviral cytokines, features that may control viral replication and prevent recurrent severe infections. Analysis of CD4+ T cell responses indicate a predominant polarization to a TH1 type,1 although the roles TH17 and TH2 cytokines and cytotoxic T cells have in patients with COVID-19 acute respiratory distress syndrome needs greater clarity. Individuals with mild or asymptomatic disease are reported to exhibit robust memory T-cell responses months after COVID-19 infection. However, it is unknown whether memory T cells in the absence of detectable circulating antibodies protect against SARS-CoV-2. Thus, identification of SARS-COV-2–specific T cells or their molecular receptor footprint4 may have future utility to assess SARS-CoV-2 exposure before antibodies arise and after their decline. At present, a full understanding of T-cell contributions in the prevention of severe COVID-19 is limited by the use of different methods to identify and profile these responses and by their use largely in cross-sectional analyses of disease groups. Moreover, investigations have focused on circulating T-cell responses in acute COVID-19, often during periods of marked lymphopenia4; thus, little is known of their functional capacity in the lung and other tissues to exert an antiviral protective or cytotoxic immunopathogenic role in convalescence.
Substantial data now demonstrate the presence of preexisting T-cell immunity to SARS-CoV-2 in blood donors either prior to the COVID-19 pandemic or more recently among those without infection.1,5,6 Memory CD4+ T cells are found in higher frequencies than are CD8+ T cells, and these likely represent responses induced by previous infection with other human endemic betacoronaviruses known to cause the common cold. Such T cells can recognize known or predicted epitopes1 within the nucleocapsid (N protein) and spike structural proteins as well as the nonstructural proteins (NSPs), NSP7 and NSP13.5 SARS-CoV-2 reactive T cells are also seen in household contacts of patients infected with SARS-CoV-2, and future studies may determine if cross-reactive T cells from previous coronavirus infections have been boosted with exposure to SARS-CoV-2. The biological implications of these findings will be significant if the preexisting T cells shape the immune repertoire to SARS-CoV-2 exposure and following vaccination as well as influence the severity of COVID-19. Overall, these data suggest T cells are another level of population-level immunity against COVID-19.
Seroprevalence data (antibodies to the SARS-CoV-2 spike protein) estimate that there may be 10 times more SARS-CoV-2 infections than the number of reported cases. Thus, it is possible that 40 million to 50 million (12% to 15% of the US population) to date may have been infected with a detectable serological response to SARS-CoV-2. However, relying on population-based natural immunity, especially for populations at risk of greater disease severity, is not wise. Boosting specific neutralizing antibodies and TH1 immunity to high levels with an effective vaccine regardless of prior immune status may further protect these individuals.
COVID-19 vaccines in development designed to prevent clinical infection, disease severity, or both show the induction of an anamnestic immune response to the spike protein with a second dose7 and can generate high levels of neutralizing antibodies comparable with or greater than those seen in sera samples from patients. The induction of sufficient CD4+ follicular helper T cells and inclusion of vaccine boosts, employed for several other vaccines where circulating antibody levels are critical for protection, may be needed to maintain levels of anti–SARS-CoV-2 neutralizing antibodies. Boosting antiviral CD8+ and TH1 CD4+ T cells recognizing spike and epitopes from other conserved regions of the proteome may also be crucial in limiting replication and disease severity. SARS-COV-2 may well follow the path of previous coronaviruses and become endemic in the population as another common cold virus.
Thus, in the few “short” months since recognition of this virus, 2 keys paths to COVID-19 adaptive immunity are being unraveled and vaccines exploiting this knowledge are in rapid development.
References
1. Grifoni A, Weiskopf D, Ramirez SI, et al. Targets of T cell responses to SARS-CoV-2 coronavirus in Humans with COVID-19 disease and unexposed individuals. Cell. 2020;181(7):1489-1501.e15. doi:10.1016/j.cell.2020.05.015PubMedGoogle ScholarCrossref
2. Suthar MS, Zimmerman MG, Kauffman RC, et al. Rapid generation of neutralizing antibody responses in COVID-19 patients. Cell Rep Med. 2020;1(3):100040. Published online June 23, 2020. doi:10.1016/j.xcrm.2020.100040PubMedGoogle Scholar
3. Robbiani DF, Gaebler C, Muecksch F, et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature. Published online June 18, 2020. doi:10.1038/s41586-020-2456-9PubMedGoogle Scholar
4. Mathew D, Giles JR, Baxter AE, et al; UPenn COVID Processing Unit. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science. Published online July 15, 2020.PubMedGoogle Scholar
5. Le Bert N, Tan AT, Kunasegaran K, et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature. Published online July 15, 2020. doi:10.1038/s41586-020-2550-zPubMedGoogle Scholar
6. Braun J, Loyal L, Frentsch M, et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature. Published online July 29, 2020.PubMedGoogle Scholar
7. O’Callaghan KP, Blatz AM, Offit PA. Developing a SARS-CoV-2 vaccine at warp speed. JAMA. 2020;324(5):437-438. doi:10.1001/jama.2020.12190 ArticlePubMedGoogle ScholarCrossref
8. Corbett KS, Flynn B, Foulds KE, et al. Evaluation of the mRNA-1273 vaccine against SARS-CoV-2 in nonhuman primates. N Engl J Med. Published online July 29, 2020.PubMedGoogle Scholar
9. Callow KA, Parry HF, Sergeant M, Tyrrell DA. The time course of the immune response to experimental coronavirus infection of man. Epidemiol Infect. 1990;105(2):435-446. doi:10.1017/S0950268800048019PubMedGoogle ScholarCrossref
10. Deng W, Bao L, Liu J, et al. Primary exposure to SARS-CoV-2 protects against reinfection in rhesus macaques. Science. 2020;369(6505):818-823. Published online July 2, 2020 . doi:10.1126/science.abc5343PubMedGoogle ScholarCrossref