Vitamin D gene: CYP11A1
The role of CYP11A1 in the production of vitamin D metabolites and their role in the regulation of epidermal functions - Oct 2014
Presented at 16th Vitamin D Workshop - https://doi.org/10.1016/j.jsbmb.2013.10.01
Association Between CYP11A1 and Vitamin D Perplexity AI Dec 2025
CYP11A1, traditionally recognized for its critical role in steroid hormone synthesis, has emerged as a key enzyme in an alternative pathway of vitamin D metabolism. This novel pathway produces biologically active vitamin D metabolites with distinct properties from classical vitamin D compounds, opening new therapeutic possibilities for various health conditions.
CYP11A1 as a Vitamin D Metabolizing Enzyme
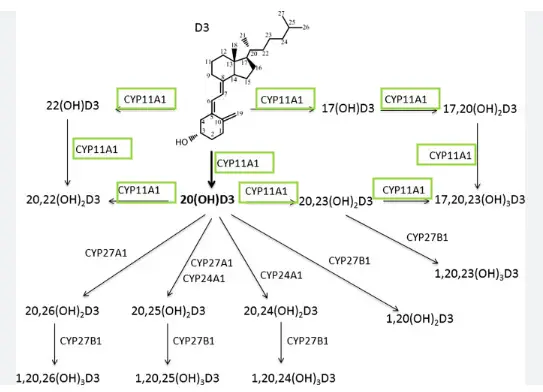
CYP11A1 (cytochrome P450 side-chain cleavage enzyme) is a mitochondrial enzyme that catalyzes the hydroxylation of vitamin D3's side chain at multiple positions, primarily at carbons 20, 22, 23, and 17, producing at least 10 different metabolites. The major product is 20S-hydroxyvitamin D3 [20(OH)D3], followed by 20,23-dihydroxyvitamin D3 [20,23(OH)₂D3], 20,22-dihydroxyvitamin D3 [20,22(OH)₂D3], 17,20-dihydroxyvitamin D3 [17,20(OH)₂D3], and 17,20,23-trihydroxyvitamin D3 [17,20,23(OH)₃D3]. This metabolic pathway represents a fundamentally different mechanism than the classical vitamin D activation pathway, as CYP11A1 hydroxylates but does not cleave the vitamin D3 side chain.[1][2][3][4]
The CYP11A1-mediated pathway operates independently of the traditional route. Importantly, CYP11A1 does not act on 25-hydroxyvitamin D3 [25(OH)D3], meaning it does not compete with CYP27B1 for activation of this major circulating form of vitamin D. This enzyme can also metabolize vitamin D2, producing 20(OH)D2 and 17,20(OH)₂D2 as major products, though with different efficiency compared to vitamin D3 metabolism.[2][4][1]
The same enzyme processes 7-dehydrocholesterol (7DHC), the precursor to vitamin D3 in skin, converting it to 7-dehydropregnenolone through side-chain cleavage. This demonstrates CYP11A1's versatility in metabolizing structurally related compounds.[4][5][6][1]
Tissue Expression and In Vivo Activity
CYP11A1 is expressed in multiple tissues beyond the classical steroidogenic organs. The placenta, adrenal glands, and epidermal keratinocytes metabolize vitamin D3 via this CYP11A1-mediated pathway, with the activity modified by CYP27B1. Evidence from ex vivo incubations of human placentae and adrenal glands with vitamin D3 has demonstrated production of 20(OH)D3, 20,23(OH)₂D3, 1,20(OH)₂D3, and 1,20,23(OH)₃D3, confirming that this pathway operates in vivo.[3][5][7][1][2]
Detection of 20(OH)D3 in human serum at levels approximately 20 times lower than 25(OH)D3 provides further evidence that CYP11A1-mediated vitamin D3 metabolism occurs endogenously in humans. In tissues with high CYP11A1 expression, the several-fold higher production of 20(OH)D3 compared to 25(OH)D3 suggests this alternative pathway may predominate locally.[8][9][1]
The enzyme's expression in skin is particularly significant, as UVB and UVC radiation—but not UVA—stimulate CYP11A1 expression when skin is incubated ex vivo or when keratinocytes are co-cultured with melanocytes. This stimulation is accompanied by upregulation of corticotropin-releasing factor (CRF), urocortin, and POMC-derived peptides. Additional regulatory factors include StAR protein, cholesterol availability, cAMP, ACTH, CRH, and cytokines such as IL-1, IL-6, and TNF-α.[10][11][12][1]
Biological Activities of CYP11A1-Derived Metabolites
The CYP11A1-derived vitamin D metabolites demonstrate remarkable biological activity through multiple mechanisms. These compounds act as biased agonists of the vitamin D receptor (VDR), expressing many but not all properties of 1,25(OH)₂D3, the classical active form. The 20S-hydroxyl group of 20(OH)D3 forms a weak hydrogen bond with His305 in the VDR ligand-binding domain. Additionally, these metabolites function as inverse agonists on RORα and RORγ receptors, which contributes to their anti-inflammatory properties.[13][14][15][16][17][18]
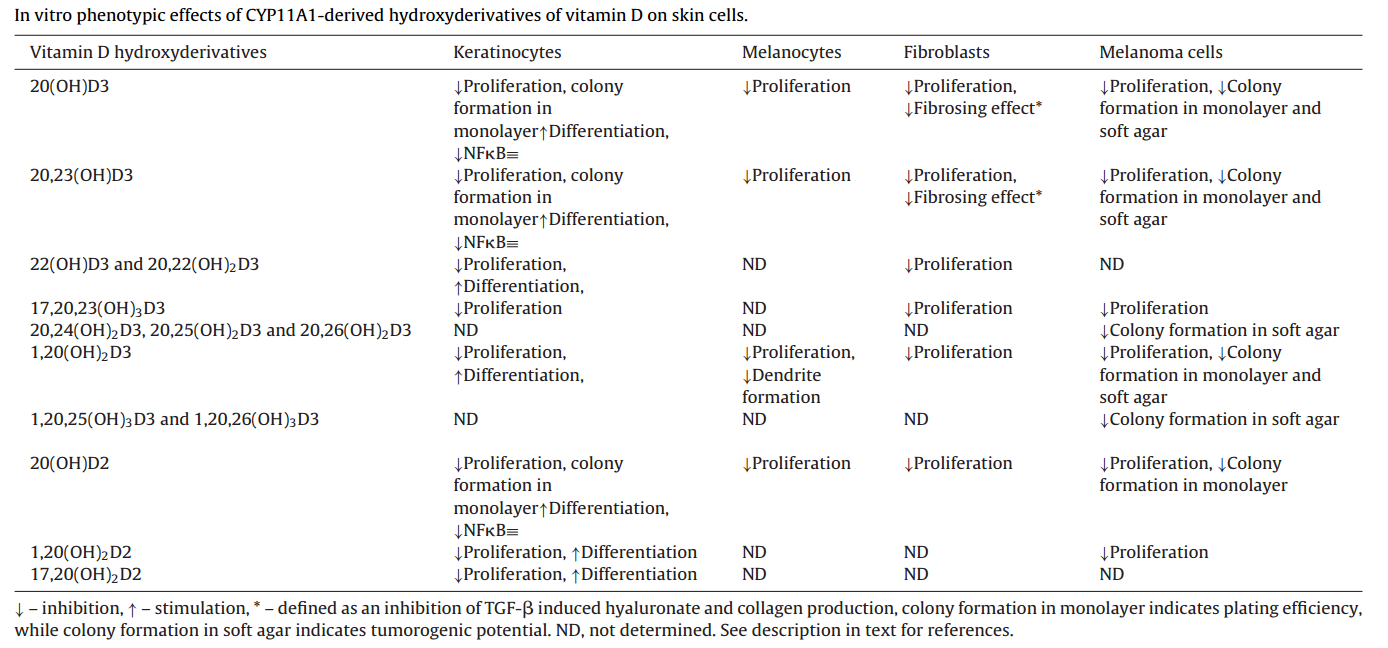
Anti-inflammatory and immunomodulatory effects are among the most significant activities. 20(OH)D3 markedly suppresses clinical signs of arthritis and joint damage in mouse models, reduces inflammatory cytokines by decreasing CD4⁺ T cells while increasing the ratio of regulatory T cells (Tregs) to CD3⁺CD4⁺ T cells. The compound decreases NF-κB activity by increasing IκBα levels and inhibiting NF-κB nuclear translocation, and inhibits production of IL-17, interferon-γ, TNF-α, and IL-2. Expression of CYP11A1 is decreased in patients with atopic dermatitis and psoriasis, suggesting it may play a protective role in inflammatory skin diseases.[19][20][21][22][23]
Antifibrotic activity has been demonstrated both in vitro and in vivo. In a bleomycin-induced mouse model of scleroderma, 20(OH)D3 exhibited excellent antifibrotic effects. The metabolites 20(OH)D3, 20,23(OH)₂D3, 20,22(OH)₂D3, and 17,20,23(OH)₃D3 inhibit proliferation of dermal fibroblasts with potency comparable to 1,25(OH)₂D3. This activity is particularly promising for treating systemic sclerosis and related fibrosing diseases, for which no effective FDA-approved treatments currently exist.[24][25]
Anticancer properties include inhibition of cell proliferation, promotion of differentiation, and reduction of tumor growth. Studies on melanoma demonstrate that 20(OH)D3 inhibits tumor growth in immunocompromised mice without visible toxicity. Classical and CYP11A1-derived vitamin D3 derivatives exhibit anticancer activities on skin cancer cell lines and inhibit GLI1 and β-catenin signaling pathways.[26][27][28][29]
Antioxidant and DNA repair functions represent another critical dimension. CYP11A1-derived metabolites protect cultured human keratinocytes and melanocytes from UVB-induced damage through activation of NRF2 and p53 signaling pathways. Vitamin D3 treatment significantly decreases DNA damage levels and increases expression of key DNA repair proteins including the MRN complex (MRE11, RAD50, NBS1), BRCA1, BRCA2, RAD51, and checkpoint kinases Chk1 and Chk2. The antioxidant mechanisms involve upregulation of glutathione reductase (GR), glutathione synthetase (GCL), superoxide dismutase (SOD), catalase, and thioredoxin reductase 1 (TXNRD1).[18][30]
Neuroprotective effects have been documented across multiple neurodegenerative conditions. In Parkinson's disease models, vitamin D pretreatment prevents neuroinflammation and loss of dopamine cells induced by oxidative stress, increases expression of C-Ret and GDNF (glial cell line-derived neurotrophic factor), and inhibits α-synuclein aggregation. Lower CYP27A1 levels in Parkinson's and multiple system atrophy patients may affect dopamine metabolism and α-synuclein aggregation through reduced vitamin D production.[31][32]
Non-Calcemic Therapeutic Potential
A defining characteristic of CYP11A1-derived vitamin D metabolites is their lack of calcemic activity at pharmacological doses. While 1,25(OH)₂D3 produces hypercalcemia at doses of 2 μg/kg, 20(OH)D3 and 20,23(OH)₂D3 are noncalcemic at doses as high as 30-60 μg/kg in rodents—approximately 300-600 times higher than doses of 1,25(OH)₂D3 that cause hypercalcemia. This remarkable safety profile makes these compounds excellent candidates for therapeutic development, as calcemic toxicity severely limits the clinical application of classical vitamin D compounds.[28][24]
The noncalcemic properties combined with potent anti-inflammatory, antifibrotic, anticancer, and neuroprotective activities position CYP11A1-derived metabolites as promising therapeutic agents for autoimmune diseases, fibrosing conditions, cancer, and neurodegenerative disorders.[33][24][28][18]
Interaction with Other Metabolic Pathways
Several CYP11A1-derived products can be further 1α-hydroxylated by CYP27B1, although with lower catalytic efficiency than 25(OH)D3. Despite lower efficiency, these 1α-hydroxylations occur in vivo, with 1,20(OH)₂D3 and 1,20,23(OH)₃D3 identified in ex utero placental and adrenal incubations. The 1α-hydroxylated derivatives show much stronger ability to activate VDR compared to their parent compounds, as demonstrated in VDRE-luciferase reporter assays and CYP24A1 expression measurements.[34][14][1]
Additionally, CYP11A1 can hydroxylate the prodrug 1α(OH)D3 to the biologically active 1,20(OH)₂D3, providing an alternative route to 25-hydroxylation for activation. The metabolites are also substrates for CYP24A1, though they demonstrate improved metabolic stability compared to 1,25(OH)₂D3.[14][1]
Clinical and Research Implications
The discovery of CYP11A1 as a vitamin D-metabolizing enzyme has expanded understanding of vitamin D biology beyond calcium and bone metabolism. This pathway represents an endogenous mechanism for producing bioactive secosteroids with therapeutic potential that extends far beyond the classical vitamin D hormone system.[33]
Current research focuses on developing analytical methods to measure circulating levels of CYP11A1-derived metabolites, which could provide new insights into vitamin D metabolite pathways and their roles in health and disease. Studies examining genetic variants in CYP11A1 have identified associations with aromatase inhibitor-related bone loss in breast cancer patients, suggesting the enzyme may have local functions in bone metabolism regulation.[9][35]
The CYP11A1 pathway also offers mechanistic insights into protective effects of vitamin D supplementation during pregnancy. In trophoblast cells, CYP11A1 upregulation can trigger excessive mitochondrial biosynthesis, oxidative stress, and inflammation that impairs neuronal stem cells. Vitamin D3 supplementation ameliorates these effects through competitive binding to P450scc and through the anti-inflammatory and antioxidant properties of its product 20(OH)D3.[36][37]
Understanding the differential regulation of cholesterol versus vitamin D3 metabolism by CYP11A1 has revealed substrate-specific protein-protein interactions with its redox partner adrenodoxin, opening possibilities for developing selective modulators of these distinct pathways.[38]
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89